Review Article
Relationships of Species of Helicotylenchus with Taxonomic Groupings of Host Plants
YS Rathore1, SN Tiwari2*
1Indian Institute of Pulses Research, Kanpur-208024/MIG 110, Green Belt, Indira Nagar, Kanpur-208026 (U.P.), India
2Department of Entomology, G.B. Pant University of Agriculture and Technology, Pantnagar ( Uttarakhand), India
Corresponding author: SN Tiwari, Department of Entomology, G.B. Pant University of Agriculture and Technology,Pantnagar ( Uttarakhand), India; E-mail: aghalenduka@yahoo.com
Citation: Rathore YS, Tiwari SN. Relationships of Species of Helicotylenchus with Taxonomic Groupings of Host Plants. J Plant Sci Res. 2016;3(1): 145.
Copyright © 2016 Rathore YS, et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Plant Science & Research | ISSN: 2349-2805 | Volume: 3, Issue: 1
Submission: 06/01/2016; Accepted: 14/03/2016; Published: 22/03/2016
Abstract
Spiral nematode, Helicotylenchus (Tylenchida : Hoplolaimidae), mostly act as ectoparasite on diverse group of plants. Studies on 33 species of spiral nematodes revealed that 7 species were host specific in monocotyledons, 6 in dicotyledons-lignosae and 5 in dicotyledons-herbaceae, and were designated as monophagous. H. digonicus, H. dihystera, H. microlobus H. multicinctus and H. psedorobustus parasitize large number of host species and were classified as polyphagous. Host affinity to taxonomic groups was species specific. Nevertheless, a general preference was more towards dicotyledons. Some species were able to parasitize host species from most primitive families to advanced ones and some included gymnosperms also in their host range. This study will help in selecting crops in particular nich or endemic areas and contribute in decision making process for their management.
Keywords: Helicotylenchus; Spiral nematode; Host Range; Dicotyledons; Monocotyledons; Lignosae; Herbaceae; Calyciferae; Corolliferae; Glumiflorae; Angiosperms; Gymnosperms
Introduction
Species of Helicotylenchus (Tylenchida: Hoplolaimidae) aregenerally known as spiral nematodes because of their coiled habit[1]. They are globally distributed and associated with root systems ofdiverse groups of plants [2]. Most are ectoparasite and feed on rootepidermis while others live half buried in the root tissues, and otherpenetrate root and live inside [3]. Information on damage potentialis not available for many species to implicate them as serious pests,however, certain species are cosmopolitan and adversely affect theplant growth and cause considerable reduction in yield. Thorsteinsonhypothesized that host selection in insects is associated with certaintaxonomic groups of plants [4]. Rathore and Lal observed that hostrange of pod borer, Maruca vitrata, is strictly restricted to plantsbelonging to leguminous group [5]. Further, Rathore and Tiwarireported that whitefly, Bemisia tabaci, preferred certain group ofplants and in some cases evolutionary lineage were seen [6]. Recent studies of Rathore and Ali and Rathore and Tiwari clearly revealedsuch taxonomic associations in Meloidogyne incognita and 63 speciesof Heterodera, respectively [7,8]. In the present review an attempt hasbeen made to analyze the relationship of Helicotylenchus with its hostspectrum.
The review encompasses thirty-three species of Helicotylenchusthe host range of which was collected from different sources [9,10].Taxonomic classification of Hutchinson was followed to analyze therelationships of different species of Helicotylenchus vis-Ã -vis hostplants [11]. In this classification, angiosperms have been divided intwo sub-phyllum- dicotyledons and monocotyledons which havebeen further classified in divisions viz., dicotyledons into lignosae(fundamentally woody plants) and herbaceae (fundamentallyherbaceous plants), and monocotyledons into calyciferae (calyxbearer-with distinct calyx and corolla), corolliferae (calyx and corollaare more or less similar) and glumiflorae (perianth is much reduced or represented by lodicules). Plant species reported as hosts of differentspecies of Helicotylenchus were aligned with families and ordersfollowing the system of Hutchinson [11]. The information thus faravailable on the clades of this nematode species was also discussed[2]. The terms monophagous (feed on plants under single genus),oligophagous (feed on plants of different genera in a single family)and polyphagous (feeding wide range of plants under different plantfamilies) were adopted as described by Bernays and Chapman [12].
Host Preference of Helicotylenchus
Information across the species elucidate that Helicotylenchus is capable to parasitize 585 plant species and out of these around 69%are dicotyledons and rest monocotyledons. Lignosae (fundamentallywoody plants) harbor 51.62% host species and 17.60% fall inherbaceae division. In monocotyledons, glumiflorae (14.36%) hasmore parasitized plants than corolliferae (10.77%) and calyciferae(5.64%). Number of families and orders follow the same trend as inthe case of host species except in glumiflorae (Table 1).
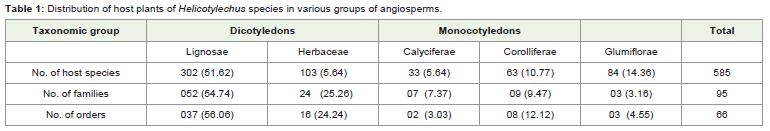
Table 1: Distribution of host plants of Helicotylechus species in various groups of angiosperms.
The three most primitive plant families viz., Magnoliaceae,Annnonaceae and Lauraceae are parasitized by 5 species only. Forexample, Magnoliaceae (Magnolales) is infested by H. dihystera and H. digonicus: Annonaceae (Annonales) by H. anhelicus, H.concavus, H. dihystera, and Lauraceae (Laurales) by H. erythrinaeand H. dihystera. On the other hand, H. dihystera parasitizes all thethree families. Ceretain less common families like Tamaraceae andJuncaceae are parasitized by H. californicus only. Similarly, hostfamilies viz., Amaranthaceae, Clusiaceae and Pedaliaceae are infestedby H. multicinctus: Typhaceae, Pittosporaceae and Vacciniaceae byH. pseudorobustus: Paeoniaceae and Punicaceae by H. erythrinae, andso on (Table 2).
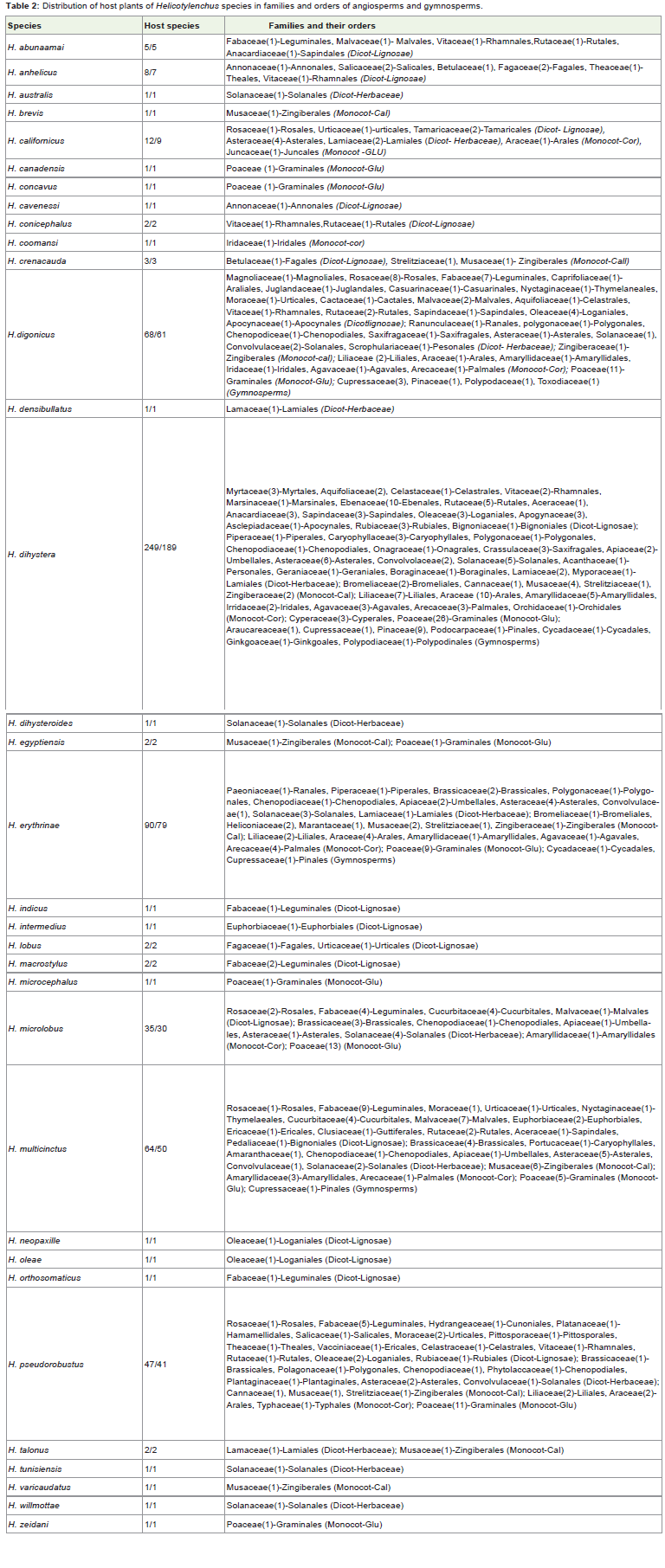
Table 2: Distribution of host plants of Helicotylenchus species in families and orders of angiosperms and gymnosperms.
The most preferred plant family is Poaceae which is parasitizedby 11 Helicotylenchus species followed by Fabaceae 10, Solanaceaeand Musaceae 9 each, and Asteraceae, Rosaceae and Rutaceae eachsharing 7 species. These plant families also harbor maximum numberof host plants (Table 3) which indicate towards diversified feeding behavior of this nematode.
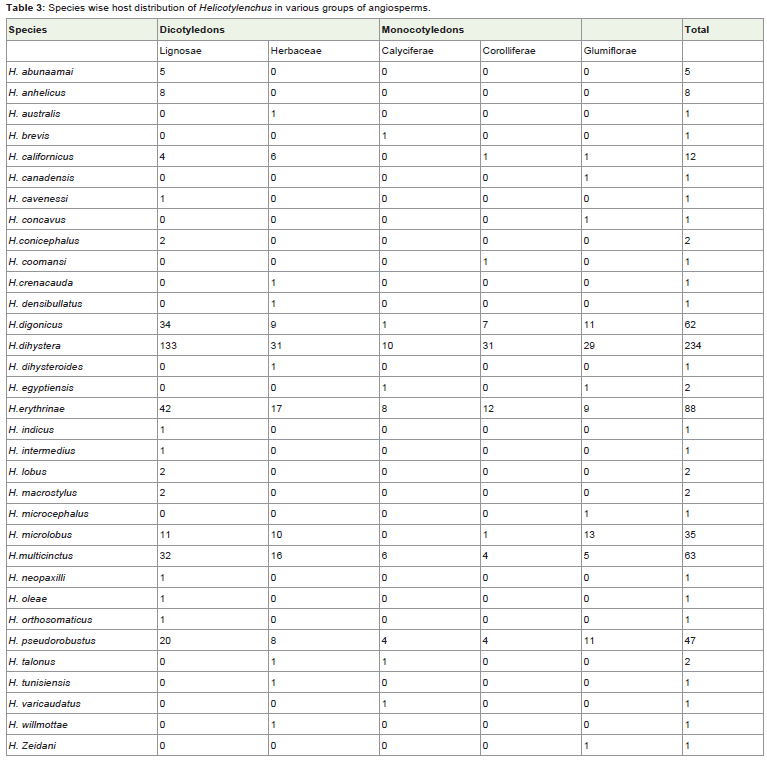
Table 3: Species wise host distribution of Helicotylenchus in various groups of angiosperms.
Species specific affiliation to taxonomic groups andphylogenetic relationships
Species specific nature provides more diversified and differentview. Among monocots, 7 species of Helicotylenchus (Table 2, Figure 1) are host specific. Helicotylenchus brevis and H. varicaudatusparasitize plants from musaceae: H. coomansi from Iridaceae, andH. canadensis, H. concavus, H. microcephalus and H. zeidani fromPoaceae only. Similarly, in dicotyledons-lignosae, H. cavenessiparasitizes plants from Annonaceae; H. indicus and H. orthosomaticusfrom Fabaceae; H. intermedius from Euphorbiaceae, and H. neopaxilliand H. oleae from Oleaceae only. In herbceae 5 species are hostspecific. Out of these, H. densibullatus was parasitic on Lamiaceae,and H. australis, H. dihysteroides, H. tunisiensis and H. willmottae onSolanaceae. Since all the above mentioned species are host specific and feed on single species of a given family, they are categorized asmonophagous.
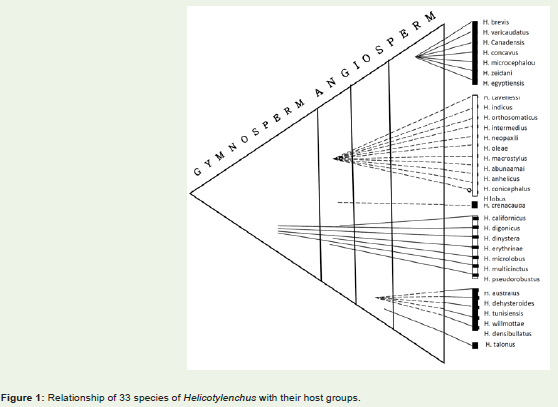
Figure 1: Relationship of 33 species of Helicotylenchus with their host groups.
One species H. macrostylus is considered as oligophagous andothers as polyphagous as they parasitize on plants from more thanone genera of different families to many genera in different familiesand orders (Table 3).
Among the polyphagous species H. californicus, H. microlobusand H. pseudorobustus are inclined more towards dicotyledons astheir host species are spread in 5, 9 and 21 families, respectively (Table 3, Figure 1). On evolutionay scale, host plants of the three species are distantly located.
Helicotylenchus dihystera has large number of host speciesranging from gymnosperm to all the divisions of dicots and monocotsin angiosperms. Among gymnosperms, it parasitizes Cycus sp.(Cycadaceae), Ginkgo sp. (Ginkgoaceae), Araucaria sp. (Araucaceae),Cupressus sp. (Cupressaceae), Abies sp., Pinus clausa, P. cubiensis,P. echidna, P. elliottis, P. paulastris, P. taeda, P. rigida, pinus sp.(Pinaceae), Podocarpus sp. (Podocarpaceae) and Polypodiaceae sp.(Polypodiaceae). Among the angiosperms, most primitive familiesMagnoliaceae (Magnoliales), Annonaceae (Annonales) and Lauraceae(Laurales) are parasitized by this nematode species. It is believedthat ancestors of angiosperms diverged from unknown group ofgymnosperms during late Triassic (245-202 million years ago) period[13,14]. Combined 3-and 4-gene rooted analysis by Bowe et al. revealthat cycads separated first from gymnosperm followed by Ginkgo andthen (Gnetales + Pinaceae) sister to a monophyletic group with allother conifer families [15]. It also implies many similarities betweengnetophytes and angiosperms. Hutchinson opined that derivation offlower structure from foliage leaves in Magnoliaceae is like the leaves and sporophylls in the more ancient gymnosperms, Pinaceae, Cycads,etc [11]. Therefore, any gymnosperm plant on which H. dihysterafeeds upon could be the progenitor of angiosperms and host speciesbelonging to Magnoliaceae (primitive of fundamentally woodyflowering plants) evolved from them. Two phylogenetic affiliationsare evident with 4 taxonomic groupings which are as follows:
1.Gymosperms--Magnoliales--Delleniales--Rosales-- Hamamelidales--Fagales--Jugandales, Urticales
2.Gymnosperm-Magnoliales-Delleniales-Bixales-Tiliales- Malvales-Euphorbiales-Rhamnales-Myrsinales-Ebenales
However, in these two evolutionary lineages, no host specieshas been reported on Delleniales in the first, and on Dellenialesand Bixales in the second. Delleniales are remotely related to theMagnoliales and show a connecting link between Magnoliales andRosales, Bixales, etc. [11].
The taxonomic groups such as i) Rosales-Leguminales, ii)Rosales-Cunoniales-Araliales, salicales, iii) Theales-Ericales,and Loganiales-Rubiales, Apocynales, Bignoniales show closerelationship and contribute a lot of host species.
In herbceae, the lineages are i) Saxifragales-Solanales-Personales, ii) Saxifragales-Caryophyllales-Onagrales,Chenopodiales, Polygonales. In monocots, order Liliales is a prolificstock and gave rise to several orders directly like Graminales,Iridales, Arales, and Agavales-Palmales. In dicotyledons-lignosae,host plants from diverse orders are selected, whereas in herbaceaesome orders like Brassicales and its progenitors and descendentsare totally rejected. Similarly, in monocots the orders evolved fromAlismatales (an ancient group, corresponding to Ranunculaceae) arenot parasitized.
Helicotylenchus digonicus parasitizes 6 species of gymnosperms(Cupressus sp., Juniperus sp., Thuja sp., Pinus sp., Sequoia sp. (Pinales)and Polypodiaceae sp. (Polypodiales) and 62 species of angiosperm.Thirty-four species belong to dicotyledons-lignosae, 9 dicotyledonsherbaceaeand 19 monocotyledons (1 calyciferae, 7 corolliferae and11 glumiflorae) embracing a total of 55 genera, 32 families and 31orders. Parasitized host species, both in lignosae and herbaceae, arefrom most primitive families to advanced families in their respectiveorders. Families Rosaceae, Fabaceae and Poaceae dominate the hostrange. Herbaceae starts with Paeoniaceae (Ranales) as a parallelismof the Magnoliaceae (Magnoliales) of dicotyledons [11]. Therefore,gymnosperms are proginator of this group also. Hence theevolutionary lineage was observed as:
Gymnosperms-Ranales-Saxifragales-Solanales-Personales.
In monocots, Liliales serve as basal stock and give rise to 6 ordersmore or less directly. No host except in Zingiberales is parasitized inany other order evolved from Commelinales and Alismatales.
Helicotylenchus erythrinae is also is a polyphagous speciesand feeds on Cycus sp. and Juniperus sp. (Cupressaceae-Pinale)in gymnosperms. It parasitizes 88 host species from 77 genera in angiosperms encompassing 23, 10, 6, 5 and 1 families in lignosae,herbaceae, calyciferae, corolliferae and glumiflorae, respectively,of dicots and monocots divisions. In both the divisions of dicots,infestation start from primitive families like Lauraceae (Laurales) inlignosae and Paeoniaceae (Ranales) in herbaceae which continue upto advanced groups. In lignosae orders viz., Rosales, Leguminales,Araliales, Salicales, Hamamelidales, Fagales, Jugandales and Urticalesevolved from Rosaceous stock and Loganiales, Rubiales, Myrtales,Rutales, Celastrales and Sapindales from different taxonomic groupscontributed for host range. In herbaceae also varied groups arepreferred. Liliales stock dominate in monocots embracing 5 familiesin 5 orders, whereas Zingiberales alone contributed host species from5 families. Family Poaceae has more parasitized plants than others.
In case of H. multicinctus no evolutionary or phylogenrticrelationships are prominent. However, it preferred host species fromvarious orders and showed close affinity with dicotyledons as well asmonocotyledons.
Investigation on phylogenetic relationships within Helicotyenchusby Subbotin et al. revealed a lot of variation within the species anddesignated them as species types [2]. They recognized 9 clades ofHelicotylenchus. Out of the species described here, H. pseudorobustuswith its species types occupied Clade I; H. digonicus Clade I, V andIX; H. dihystera Clade II; H. multicinctus Clade III, and H. brevisClade VII. Fortuner et al. opined H. rotundicauda as a synonym ofH. dihystera [16]. Subbotin et al. also support the synonymy of H.rotundicauda with H. dihystera [2]. Samples of H. dihystera fromWest Africa (Sawadogo et al, 2009) clustered in Clade iii with H.muticinctus [17]. According to Bae et al. the aforesaid samples alsoclustered with H. microlobus. Sher synonymized H. microlobus withH. pseudorobustus [18]. On host front, H. dihystera, H. digonicus, H.microlobus, H. multicinctus and H. pseudorobustus are polyphagousand predominately prefer dicotyledonous plants. Taxonomicgroupings also support their synonymy. H. brevis in Clade VII ismonophagous and parasitize monocotyledons. It does not share anyother Helicotylenchus species in this clade.
The information provided in preceding paragraphs on taxonomicgroupings of host species has predictive value. In endemic areas, thiswill help in selection of appropriate varieties, formulation of bettercrop rotations, execution of important agronomic practices andadoption of better nematode management practices.
Acknowledgements
Authors are grateful to Mr. Akhilesh Rathore for extending allpossible help in retrieving the information.
References
- FAO (2005) FAO year book, Rome Italy.
- NAERLS (2009) Annual Agric performance survey Report of Nigeria. National Report 2009.
- IITA (2010) International Institute of Tropical Agriculture (2010).Annual Report, IITA Ibadan, Nigeria.
- Onwueme IC (1978) Tropical root and tuber crops, Cassava, Sweet potato Cocoyam. John Wiley and sons. N.Y 224pp.
- Asiedu R, Wanyera NM, Ng SYC, Ng NQ (1997) Yams In: Biodiversty in Trust. D. Fuccillo, L.Sear & P.Stapleton, Nematrropica 1: 39-40.
- Emehute JK, Ikotun UT, Nwauzor EC, Nwokocha HN (1997) Crop protection. In: Food yams; Advances in Research. GC Orkwor, R Asiedu, IJ Ekanayake. (ed). NRCRI /Nigeria. PP143-186.
- Onyenobi FI (1992) Root knot nematode Meloidogyne. An economic Problem in Nigeria. In: the biology and control of nematode of food crops in Nigeria. Pp 37-48.
- Onyenobi FI, Aghale DN, Okorocha EA (2006) Post -harvest studies of yam pre-treated with bio and synthetic pesticides to control Meloidogyne incognita at cultivation. Proceedings of the 40th Conference of the Agriculture Society of Nigeria, Abia Pg 463-466.
- Aghale DN (2011) Evaluation of Botanicals in the control of Root Knot nematode M. incognita (Kofoid& Whitehead, 1919). Ph.D Thesis, Dept of Crop Production/Protection, Abia State University, Uturu, Nigeria.
- Aghale DN, Okorocha A, Njoku, Nwanyieze (2010) Evaluation of Different Botanicals for root knot nematode control. Journal of Agric Biology & Food Research 3: 23 -240.
- Aghale DN (2011) Efficacy of crude plant extract at different rates of application in the control of root knot nematode (M. incognita) in yam production. International Journal of Agric food & Ecosystem Research 3: 200-215.
- Husssey RS, Barker KB (1973) “A comparison of methods of collecting inocula of Meloidogyne spp., including a new techniqueâ€. Pl Dis Reptr 57: 1025-1028.
- AOAC (1990) Official method of Analysis. 15th Edition, Association of official Analytical Chemist, Washington D.C USA.
- Arora SK (1981) Analytical Technique for quality Evaluation. Forage Research 7: 196-213.
- Nwauzor EC, Fawole B (1985) Root knot nematode Meloidogynesppon edible yam Dioscoreaspp in Nigeria. Effect on tuber yield. 15th Ann Conf. of NSPP, NRCRI, Umudike, March 3-8.
- National Root Crops Research Institute (2005) Annual report, 2005. 260pp.
- SAS (1999) Statistical Analysis System. The SAS system for Windows version 8. SAS Institute Inc. Cary, NC, USA.