Research Article
Seed the Source of Systemic Botrytis cinerea in Lettuce (Lactuca sativa L.)
Yahaya SM* and Fagwalawa LD
Department of Biology, Kano University of Sciences and Technology, Wudil -Kano State P.M.B. 3244, Nigeria
Corresponding author: Yahaya SM, Department of Biology, Kano University of Sciences and Technology, Wudil -Kano State P.M.B. 3244, Nigeria; E-mail: sanimyahya@yahoo.com
Citation: SM Y, LD F. Seed the Source of Systemic Botrytis cinerea in Lettuce (Lactuca sativa L.). J Plant Sci Res. 2015;2(2): 141.
Copyright © 2015 Yahaya SM, et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Plant Science & Research | ISSN: 2349-2805 | Volume: 2, Issue: 2
Submission: 27/10/2015; Accepted: 30/11/2015; Published: 05/12/2015
Abstract
Studying the epidemiology of the wide spread of necrotrophic pathogen Botrytis cinerea is of economic importance in the production and seed export of lettuce and other horticultural crops. This study showed that seed is the prime source of systemic B. cinerea in lettuce plants. The presence of the pathogen in plants grown from fungicide treated seeds suggests that the pathogen is systemic. Infection arises from both internal and external seed infection. This conclusion is based on detection of seed infection from plants which were grown in controlled conditions from infected seed collected from flower inoculatedplants. Fragment analysis found that 62.5% of seed isolates detected from plants which were grown from non-fungicide treated seeds was identical to the inoculation isolate. In contrast, all seed isolates detected from plants which were grown from fungicide treated seed have different genotypes to the inoculating isolate. The results showed that with time as the plant grew, the infection from the seed spread to root, stems and leaves systemically without exhibiting symptoms. Thus seed infection spread to other parts of the plants is age dependent. This confirmed the systemic spread of the seed isolate, but showed that other B. cinerea isolates were present and that single plants can host multiple isolates. This shows that the endophytic phase is an important component ofthe population dynamics of B. cinerea.
Keywords: Botrytis cinerea; Endophyte; Lactuca sativa; Systemic infection
Introduction
Botrytis cinerea an Ascomycetes, family Sclerotiniaceae is as a model necrotic pathogen which grows endophytically withoutshowing symptoms in greenhouse and outdoor crops, resultingin great economic losses [1-3]. Elad et al. reported that B. cinereacan enter into the host body and remain dormant or quiescent for avarying periods resulting in a symptomless endophytic infection [4].However, during quiescence, B. cinerea is able to grow endophyticallyby avoiding the induction of plant defences by not producing toxiccompounds or by producing toxic compounds at a level which allowsit to continue to grow in the presence of host plant responses withoutbeing detected [3,5,6].
Fully endophytic B. cinerea, like other endophytic fungi, areonly disseminated by seed; being highly and efficiently dispersed by their host seed and so they need not have spores as a means ofdissemination [7,8]. With the start of seed production, the endophytegrows upward in the plant and infects the outer layers of the seed;these transfer endophytes from the plant batch to the seed, [9,1,4,6).However, seed infection is only important in circumstances wherethe pathogen can spread into the plant [9]. Elias et al. reported thathighest rate of infection of B. cinerea occurred in the roots of plantsgrown from infected seeds, but as the plants grew the infectionmoved upwards into newly produced tissues [6]. The general trendis that infections progress from the roots, through the stems to theleaves and finally into the seed without causing symptoms [10,11,6].Generally, internal infection may depend on which part of the floweris infected thus affecting the likelihood of the pathogen transferringinto the seed.
Although endophytes are usually symptomless they can produce effects ranging from beneficial to pathogenic [12-15]. However, thebenefits conferred by these fungi is dependent on host species, hostgenotype and environmental conditions [16,17]. One of the benefitsto the host is the increase in host shoot and/ or root biomass, usuallydue to the induction of plant hormones by the fungus. Other benefitsinclude endophyte-induced changes in morphology and physiologyleading to an increase in vigour, survival and resistance to droughttolerance. The increased growth seen in endophyte infected plantscould be due to the production of growth hormones such as indole-3-acetic acid (IAA) and cytokinins or the enhancement of host nutrientuptake ability, especially of phosphorus and nitrogen. Some systemicendophytic fungi are able to produce physiologically active alkaloidsin host plant tissues, providing a defence against herbivorous [18,17].
The harmful effects of endophytes in plants include preventingthe flowering of their plant host. For example with Botrytis anthophilaattacking red clover, its spores may replace the pollen grains on theanther of the host plant and giving a characteristic ash-grey colourto the pollen. The spores are transmitted by bees and germinate withpollen grains on the stigmas of healthy plants, usually leading to thedevelopment of an intraseminal mycelium from which the systemicinfection of the adult plant is derived. The diseases have been foundto be widely distributed and have adverse effects on the fertility of redclover [19].
However, the endophytic colonisation of plants by B. cinerealead to, systemic infection of the host [18]. Therefore the endophyticgrowth of B. cinerea has become of great concerned for the foodproduction industry as it results in serious unexpected post-harvestproblems due to the decay of the produce without any prior warning.Therefore in order to increase availability and to reduce the cost ofagricultural produce it is important to study the systemic endophyticspread of B. cinerea which will provide information in designing ofcontrol strategies. Thus to understand the epidemiology, we testedthree hypotheses. First if seed borne endophytic B. cinerearesultsin the establishment of internal infection in lettuce plants. Secondinfection from root spread to the upper parts of the plants in lettuce.Third plant age in important in the spread of internal infection.
Material and Methods
Botrytis cinerea culture
Three types of pumpkin seed varieties (5 kg for each variety) werestudied, two of which were planted in Egypt (kafr saad and kafr elbatikh),and the third Chinese variety namly Hongli was planted inChina. Chinese variety was obtained from the local market in Cairowhereas kafr saad and kafr el-batikh were obtained from their regions.The seeds were cleaned from foreign materials, hand dehulled and theobtained kernels were ground by using electrical grinder and used foranalysis. All reagents were purchased from Sigma-Aldrich.
Analytical Method
B. cinerea isolate, B1.1 (isolated at Reading in 2005) was maintained on Botrytis selective medium (BSM) before being transferred to 3% malt extract agar (MEA CM0059, Oxoid, Basingstoke). After theculture covered approximately 1/3 of each plate, they were exposedto continuous UV-light to encourage sporulation. Botrytis selectivemedia was used in the detection and isolation of the fungal pathogenfrom plant tissues.
Seed stock
The seed used in this experiment was collected from flowerinoculated lettuce plants which were initially grown from lettuceseed (Tom Thumb variety, Fothergills Seeds, Newmarket), whichwere harvested in 2009. The seed was sterilized by soaking in 100mlof (0.1g/l) the systemic fungicide Shirlan (active ingredient 500g/lFluazinam, Sygenta Crop Protection UK limited) for 2h and drying over night before sowing.
Plant growth and inoculation
Infected seed from flower inoculated plants were divided into twotreatments one was treated with the fungicide Fluazinam followingthe same method used to sterilize the initial seed stock. Fungicide andnon-fungicide treated seeds were separately grown each in twenty,15cm pots, each with four seedlings per pot. Pots were arranged intwo blocks, each consisting of 20 pots (10 pots each for fungicideand non- fungicide treated seeds). At the two leaf stage, Block 1 wasselected and inoculated with fungal spores by gently tapping theback of Petri plates containing a 14-day-old sporulating culture of B.cinerea B1.1. The seedlings were left for 10 min to allow spores to settle, then transferred into black polyethylene bags (Figure 1) and stored in the laboratory under ambient conditions (to prevent overheatingand condensation) for 24h before removing the polyethylene bag.Seedlings were reared in a controlled environmental room (22 °C +/-,2 °C, RH 66).

Figure 1: Seedlings left for 24h in a black polythene bag.
Isolation of Botrytis cinerea from letce seedlings
The experiment was performed twice (April/May 2011 and June/July, 2011). In each case, one week after inoculation one seedling wascollected from each of the 40 pots in the two blocks and divided intoone cm-long sections of secondary root, 1 cm diameter leaf discsand hand-cut 1 mm sections of stems. These were sterilized andindividually plated in BSM plates. This was repeated at intervals ofthree, six and eight weeks after inoculation. The growth of B. cinereafrom the seedling sections after each plating test was counted andrecorded.
Genetic characterization of B. cinereaisolate (microsatellite)
Extract of DNA from sections of roots, stems and leaves isolatedeight weeks after inoculation. Mycelium (100mg) was scratchedfrom Petri plates and manually ground in a pestle and mortar in thepresence of liquid nitrogen. The resulting fine powder was transferredinto a 200 ml effendorf tube and DNA was extracted using the DN easyPlant Mini Kit (Qiagen, West Sussex, UK), stored at -20 °C and theconcentration of the DNA as adjusted to 10ng/μl using a NanodropND 1000 Spectrophotometer (Applied Biosystems UK). The B.cinerea DNA isolates were characterised using nine microsatellitemarkers designed by Fournier et al. [20] which were labelled withFAM (Blue), HEX (Green) and NED (Yellow). Each reaction mixturecontained 25 μl of Biomix (Bioline, UK), 20 μl of water, 1.5 μl of eachforward and reverse primer and 2 μl of DNA template. The PCR ofthe SSR was repeated several times for successful amplification. Theproducts from successful amplifications were multiplexed in threecombinations; (BC1, BC4, BC9), (BC3, BC6, BC10), (BC2, BC5,BC7). These groups were chosen to avoid overlapping the allele sizeranges of the primers in each mix. The PCR was run with an initialdenaturing step of two minutes at 94 °C for BC1, BC2, BC3, BC5, BC6and BC9 followed by 35 cycles of one minute at 94 °C, one minuteannealing at 53 °C, 30 seconds at 72 °C and 5 minutes at 72 °C. WhileBC4, BC7 and BC10 an annealing temperature of 59 °C was used. Theproducts were submitted for fragment analysis by Source Bioscience,UK. The results of the fragment analysis were scored by using Genemapper software (Applied Biosystem).
Experimental design
Factorial block design was used with three main factors (a) seedfungicide treatment (b) seedling inoculation and (c) time. One blockof seedlings was dry inoculated two weeks after sowing with fungalisolate B1.1. Although the density of the spore deposition was not measured, the technique was similar to the one used for the initialcommercially purchased lettuce seeds in 2009. One week afterinoculation one seedling was collected from each of the 40 pots,washed under running tap water and sterilized by spraying with70% ethanol and then allowed to air dry in a sterilized lamina flow.Seedlings were divided into roots, stems and leaves and plated inBotrytis selective media (BSM) at 18 °C for one week. The remainingseedlings were sampled and plated at interval of three, six and eightweeks after inoculation. At eight week after inoculation, the 61colonies isolated from roots, stems and leaves were subcultured andfragment analysis was performed following the previously describedprotocols.
Statistical analysis
B. cinerea colony counted at the end of the experiment (eightweek after inoculation was compared between the four differentexperimental treatments using a generalised linear model approach(GLM) in Mini tab software.
Results
Systemic and endophytic infection of B. cinerea
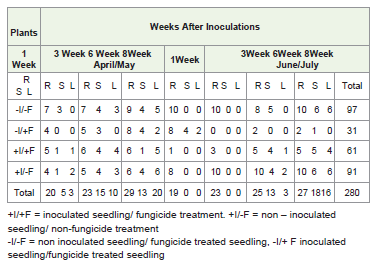
B. cinerea was recovered from the surface-sterilized sections ofhealthy leaves, stems and roots for all treatments in April/May 2011and June/July 2011, with the exception of a single sample plated oneweek after inoculation in April/May 2011, where all samples became contaminated (Table 1). Although growth of B. cinerea was recordedfrom the second, third and fourth sampling of April/May 2011, thedifference between the treatments was very close to significant (P=0.06). In June/July 2011 experiments, infection was initially mostcommon in the roots with clean stems and leaves. However, as theplants grew the infection spread into stems and then into leaves. Inthe last plating test eight weeks after inoculation colony count fromthe roots stems and leaves was relatively one. (Table 1, Figure 2, time on the spread of infection P< 0.002).
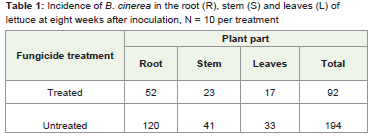
Table 1: Incidence of B. cinerea in the root (R), stem (S) and leaves (L) of lettuce at eight weeks after inoculation, N = 10 per treatment.
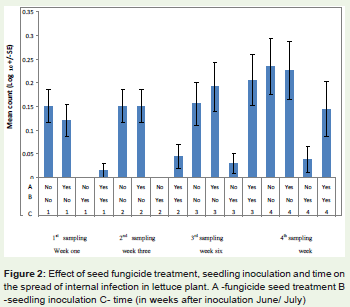
Figure 2: Effect of seed fungicide treatment, seedling inoculation and time on the spread of internal infection in lettuce plant. A -fungicide seed treatment B -seedling inoculation C- time (in weeks after inoculation June/ July).
Seed fungicide treatment
Fungicide treatment of the seed before sowing cleared most of theseed infections and greatly reduced the incidence of infections in allthe plant tissues plated at first and second plating test. At three weeksafter inoculation (Table 1, Figure 2) plants grown from fungicidetreated seed in June/July had no stem and leaf infection but moderatelevels in their roots; the stems and leaves were significantly differentfrom the ones grown from the untreated seed (F 1,319 = 32.65, P =0.001, Table 2).
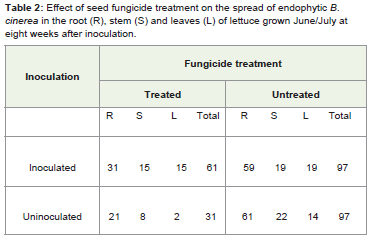
Table 2: Effect of seed fungicide treatment on the spread of endophytic B. cinerea in the root (R), stem (S) and leaves (L) of lettuce grown June/July at eight weeks after inoculation.
At the fourth plating test, eight weeks after inoculation leaves hadless infection but the roots and stems were not affected by the seedfungicide treatment. The effect of seed fungicide treatment interactedsignificantly with the effect of seedling inoculation (F 1,319= 4.76, P=0.030, Table 3).
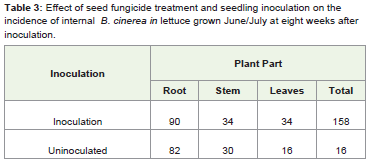
Table 3: Effect of seed fungicide treatment and seedling inoculation on the incidence of internal B. cinerea in lettuce grown June/July at eight weeks after inoculation.
However, there was no significant interaction between seedfungicide treatment, seedling inoculation and time on the spread ofinfection (F3, 19 = 0.38, P= 0.771).
Seedling inoculation
Seedling inoculation in both the April/May and June/Julyexperiments did not affect infection levels at the 1st and 3rd weeksafter inoculation, but altered infection frequency at 6th and 8th weeksafter inoculation. However, there was no significant interactionbetween seedling inoculation and time on the spread of internalinfection (Table 4, F 3,319 = 1.50, P= 0.215).
Effect of time on the spread of internal infection
The initial seed source was infected seed. All the plants whichwere grown without seed fungicide treatment initially had a high rateof root infection, but stems and leaves were free of infection in testsconducted after inoculation (Table 4). However, as the plant grew theinfection moved into stems and then leaves. There was a significantinteraction between time and location of infection (Table 4, F 3,319 =4.92, P= 0.002).
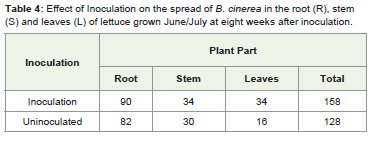
Table 4: Effect of Inoculation on the spread of B. cinerea in the root (R), stem S) and leaves (L) of lettuce grown June/July at eight weeks after inoculation.
Genotypic characterization of the isolates recovered
Isolates recovered from the roots, stems, and leaves of experimental plating in the June/July experiment eight weeks afterinoculation were genotyped at nine microsatellite loci. The isolatedetected from the initial seed used to grow the plants (haplotypecoded A), was recovered from 62.5% of the plants which were notinoculated at seedling stage and in some cases the isolate was recoveredthroughout each plant. All isolates recovered from inoculated plantshad a different genotypic identity from the isolate recovered from thesource seed.
Discussion
The study establishes that seed is the source of systemic B. cinereainfection in lettuce. Infection is initially confined to the root in plantsgrown from infected seed, and as the plant grows infection movesinto developing tissues. In April/May, 2011, when the plants weregrown from fungicide and non-fungicide treated seed, although nocolony count was recorded at one week after inoculation due to thecontamination of the samples; subsequent plating tests show thatroots carried more infection of B. cinerea. When the experimentwas repeated in June/July 2011, the distribution of B. cinerea in theparts of lettuce plant was somewhat different (Table 1). From theplating test at one and three weeks after inoculation the infection wasconfined to the roots while the stems, and leaves were infection free inplant grown from non-fungicide treated seeds. However, the platingtest at one and three weeks after inoculation shows that roots, stemsand leaves were clean in plants grown from fungicide treated seed.At six weeks after inoculation both treatments show that roots havethe highest infection rate, but there was also a movement of infectionfrom the roots to the upper parts of the plant. The plating test ateight weeks after inoculation shows a balance of infection betweenthe plant tissues. In a related experiment, Elias et al. [6] found thathealthy looking lettuce plants without any visible disease symptomsmay harbour live B. cinerea in their roots, stems and leaves. Barnesand Shaw [9] found that Primula x Polyantha plants grown frominfected commercial seeds had a high level of root infection. Since theplants were grown in isolation propagator, they concluded that theinfection in the roots was likely to have spread into the plant.
In this study it is established that as plants grow, B. cinerea infectionmoves to newly produced tissues. It is possible that B. cinerea goesinto a quiescent phase before spreading to the stems and leaves Barnesand Shaw [9]. Seed fungicide treatment was effective in removingsurface contamination. The presence of the pathogen in plants grownfrom fungicide treated seeds suggests that the pathogen is systemic.Infection arises from both internal and external seed infection (Table 3). Sowley found that seed fungicide treatments significantly lowerseedling infection rate therefore most of the infection comes fromthe seed coat [10]. Surface sterilization or seed washing confirmedthat infection was largely due to surface contamination [21,17].Kabeere et al. reported that infection of maize seeds by Fusariumsubglutinans was reduced from 96% to 8% after surface sterilization[22]. They showed that the majority of seed inoculum was on the seedsurface because there was a substantial reduction in recovery of F.subglutinans when seeds were surface sterilized.
Seedlings grown from non fungicide treated seeds tended tohave a higher spread of fungal infection as shown in Table 3. In bothApril/May 2011 and June/July2011, there was a general increase in the spread of infection from one week after inoculation to eightweeks after inoculation as shown in Table 1, in both fungicide andnon fungicide treated seedlings. The general increase in infectioncould be due to B. cinerea spreading within plant tissues withoutcausing symptoms. There are two alternative hypotheses for thepattern of infection described. First, each part of the plant couldbe independently infected. Second, the pattern in the result ofsystemic infection resulting from the initial source infection. Thesedata support the later hypothesis, given the timing of expression ofinfection and the results from the fragment analysis.
Stewart and Franicevic [23] investigation of the spread of internal infection by Botrytis species, revealed transmission of B. allii frominfected onion seed into the seedling tissue. However, the percentageof seedling infection was less than that of seed infection whichsuggests that not all seed infection results in seedling infection. Alsono differences were found in the amount of B. allii infection betweenplants grown from water-washed and fungicide treated seeds. Thissuggest that B. allii is present within the seed rather than on theseed surface. The ability of endophytic B. cinerea to be disseminatedby seed makes it the most widely distributed disease of vegetables,ornamentals, fruits, and field crops throughout the world [3].
The study show that recovery of infection in plants grown from B. cinerea infected seed varies with time the plant age. Azevedo andWelty (1995) found that the recovery of Acremonium coenophiilumfrom infected plants varies with seedling age, root type andmedium. Similarly, Kelly and Wallin found that Aspergillus flavus isconfined to the roots of maize at the 3-leaf stage while by the 4-leafstage it is uniformly distributed in the root, stem and leaves [24].Aspergillus flavus became confined to the root and lower portionsof the stem at the 5-leaf stage. In Primular x polyantha grown frominfected commercial seeds, Barnes and Shaw [9] found the highestconcentration of B. cinerea infection in the root which later spreadto other parts of the plant endophytically with symptoms appearingthree months later at flowering. Botrytis fabae was isolated from allparts of symptomless field bean seedlings grown from infected seeds.However, whether plants were likely to show infection was found tobe temperature dependent [25].
The SSR results shows that isolates recovered from roots, stemsand leaves of 62.5% of uninoculated plants was similar to thegenotype of B. cinerea from the initial seed source. Out of that 6.4%of uninoculated plants have isolates in their roots, stems and leavessimilar to the B. cinerea isolated from the source seed. Therefore,Isolates from the inoculated plants have different genotypic identitiesfrom that of the seed source (Table 5).
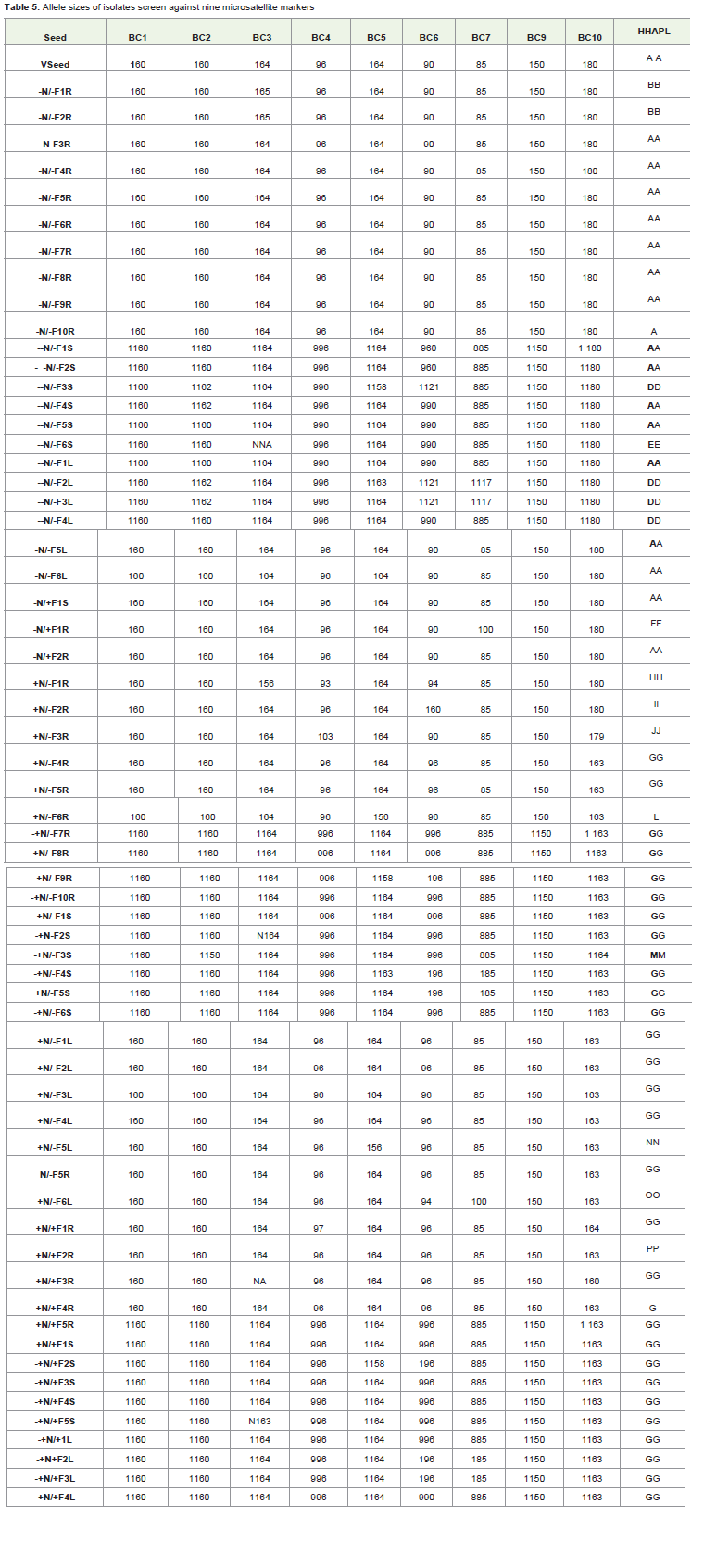
Table 5: Allele sizes of isolates screen against nine microsatellite markers
To conclude, this study has established that seed is the sourceof infection by B. cinerea in lettuce seedlings and as the plant growsinfection spreads to newly produced tissues leading to a long-livedsystemic endophytic infection in all parts. The finding here aresimilar to the results of Sowley [10] and; Shafia [11] that susceptibilityof lettuce plant to infection increases with aging, the incidence ofinfection on matured leaves was higher than of younger and healthyleaves. Previous works have suggested that B. cinerea is root specificbut this study suggested that while B. cinerea may be initially confinedto the roots it may eventually spread to other parts of the plant.It istherefore recommended that a good quarantine system should beput in place to check all commercial seed stocks for planting andespecially those earmarked for export and import in order to reducethe spread of systemic B. cinerea from one country to another.
Acknowledgement
Thanks to Mark, Sue, Lee, Val Jasper, Jang, Anushka and Matthewfor their support and assistance.
References
- Elad Y, Williamson B, Tudzynski P, Delen N (2004).Botrytis spp. and diseases they cause in Agricultural systems-An introduction. In Y.Elad (eds). Botrytis: Biology, Pathology and Control. 1-6 Springer, Dordrecht. The Netherland.
- Tejesvi M, Kini KR, Prakash HS, Shatty HS (2007) Genetic diversity and antifungal activity of species of pestalotiopsis isolated as endophytes from medicinal plants. Fungal Diversity 31: 37-54.
- Williamson B, Tudzynski B, Tudzynski P, Van Kan JA (2007) Botrytis cinerea: the cause of grey mould diseases. Mol Plant Pathol 8: 561-580.
- Elad Y, Zimand G, Zaqs Y, Zuriel S, Chet I (2007) Use of Trichoderma harzianum in combination or alteration with fungicides to control cucumber grey mould (Botrytis cinerea) under commercial conditions. Plant pathology 42: 324-332.
- Oses R, Valenzuela S, Freer J, Sanfuetes E, Rodriquez J (2008) Fungal endophytes in xylemof healthy Chilean trees and their possible role in early wood. Fungal Diversity 33: 77-86.
- Elias SNK, Fiona MD, Shaw MW (2010) Persistent, Symptomless, systemic and seed-borne infection of lettuce by Botrytis cinerea. Eur J Plant Pathol 126: 61-71.
- Saikkonen K, Ion D, Gyllenberg M (2002) The persistence of vertically transmitted fungi ingrass metapopulations. Proc Biol Sci 269: 1394-1403.
- Stone JK, Polishook JD, White JF (2004) Endophytic fungi. Biodiversity of fungi:inventoryand monitoring methods, Burlington, MA, USA: Elsevier, 241-270.
- Barnes SE, Shaw MW (2003) Infection of commercial hybrid seed by Botrytis cinerea and latent diseases spread through the plants. Phytopathology 93: 573-577.
- Sowley ENK (2006) Epidemiology of Botrytis cinerea. PhD: University of Reading, UK.
- Shafia A (2009) Latent infection of Botrytis cinerea. PhD thesis, School of Biological Sciences, University of Reading, UK.
- Bacon CW, White LF jr. (2000) Microbial endophytes Markel Dekker inc., New York, Basel USA: 487.
- Arnold AE, Maynard Z, Gilbert GS, Kursar TA (2000) Are tropical fungal endophytes hyperdiverse? Ecol Lett 3: 267-274.
- Stone JK, Viret O, Petrini O, Chapela I (2000) Histological studies of host penetrationand colonization by endophytic fungi. In host wall Alteration by parasitic fungi. 115-128, American Phytopathological Society Press, St Paul, MN.
- Photita W, Lumyong P, Hyde KD (2001). Endophytic fungi of wild banana (Musa acuminate) at Doi suthep pui National park, Thailand. Mycological Research 105: 1506-1514.
- Faeth SH, Sullivan TJ (2003) Mutualistic asexual endophytes in a native grass are usually parasitic. Am Nat 161: 310-325.
- Faeth SH, GardnerDR, Hayes CJ, Jani A, Wittlinger SK, et al. (2006) Temporal and spiral variation in alkaloid levels in Achnatherum robustrum, a native grass infected with the endophyte Neotyphodium. J Chem Ecol 32: 307-324.
- Tan RX, Zou WX (2001) Endophytes: a rich source of functional metabolites. National product report 18: 448-459.
- Silow RA (2009) A systemic diseases of red clover caused by Botrytis anthorphila Bond. Transaction of British Mycological Society 18: 239-248.
- Fournier E, Giraud A, Loiseau D, Vantrin A, Estoup M, et al. (2002) Characterization of nine polymorphic microsatellite loci in the fungus Botrytis cinerea (Ascomycota). Molecular Biology Ecology Notes 2: 253-255.
- Burgess DR, Bretag TW, Keane PJ (1997) Seed to seedling transmission of Botrytis cinerea chickpea and disinfestation of seed with moist heat. Aust J Experimental Agric 37: 223-229.
- Kabeere F, Hill MJ, Hampton JG (1997) The transmission of Fusarium subglutinans from maize seeds to seedlings. Aust Plant pathol 26: 126-130.
- Stewart A, Franicevic SC (1994) Infected seed as a source of inoculum for Botrytis cinerea infection of onion bulb in store. Aust Plant Pathol 23: 36-41.
- Kelly SM, Willin JR (1986) Systemic infection of maize plants by Aspergillus flavus. Aflatoxins in maize. El Batan. Mexico, CIMMYT, Mexico, DF Mexico.
- Harrison JG (1978) Role of Seed-borne infection in epidemiology of Botrytis fabae on field beans. Transactions of the British Mycological Society 70: 35-40.