Does Botrytis Cinerea Influence Parasitization of Myzus Persicae by Aphidius Colemani?
Yahaya SM*, Ali MU and Abdullahi H
Corresponding author: Sani Mohammed Yahaya, Department of Biology, Kano University of Science and TechnologyWudil, PMB 3244, Kano State, Nigeria; E-mail: sanimyahya@yahoo.com
Department of Biology, Kano University of Science and Technology Wudil, PMB 3244, Kano State, Nigeria
Citation: Yahaya SM, Ali MU, Abdullahi H. Does Botrytis Cinerea Influence Parasitization of Myzus Persicae by Aphidius Colemani? J Plant Sci Res.2015;2(2): 132.
Copyright © 2015 Yahaya SM, et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Plant Science & Research | ISSN: 2349-2805 | Volume: 2, Issue: 2
Submission: 04/08/2015; Accepted: 25/08/2015; Published: 01/09/2015
Abstract
The dynamics of the insect herbivore and its natural enemy appears to be influenced by the presence of the pathogen. More M. persicae and parasitoids Aphidius colemani were grown on lettuce plants free from B. cinerea infection. The size of M. persicae and its parasitoid, A. colemani was significantly larger when reared on uninfected plants. However, similar parasitoid sex ratio, 50:50 was found between parasitoids emerging from both infected and uninfected plants. The experiment provide further evidence that healthy plants free from B. cinerea infection provided better quality food resources for the aphid host which encourage fitness and results in larger sized offspring. Therefore the negative effect of B. cinerea on the interaction between aphid and its natural enemy parasitoid was determined in the present study may be very valuable in understanding interaction at higher trophic level and may also provide a clue for design of disease management strategies.
Keywords: Aphidius colemani; Botrytis cinerea; Myzus persicae; Lettuce plant
Introduction
Aphids are important agricultural pests, which causes direct andindirect damage to crops. The direct damage occur by aphids feedingon crops were they use their stylets for piercing into the phloem [1,2]while the in indirect damage occur when aphids act as vectors of viraldiseases resulting in significant diseases to their host plants [3,4].
Aphids are attacked by a range of natural enemies includinghymenopteran parasitoids (e.g. Aphidius species such as Aphidiuscolemani, A. matricariae, A. ervi and A. abdominalis), and a widerange of arthropod predators [5,3,6,7]. High aphid numbers on aplant serves as a resource for their natural enemies. Many studieshave investigated the potential effects of natural enemies for keepingaphid population sizes below economic injury levels [8,9].
Previous researches by Yahaya and Fellowes showed that aphidsindirectly interact with necrotrophic pathogens such as B. cinereawhen sharing host plant [10]. Where aphid and B. cinerea encounter one another on the same host plant both aphids and pathogensmay induces the host plant to release various defensive chemicals[11], these chemicals in addition also serve as cues which attractedaphid natural enemies [9]. Although there have been fewer studiesexamining the effect of necrotrophic pathogens such as B. cinereaon the interaction between aphids and their natural enemies and itspotential influence on successful biocontrol but Persson, Aquilino etal., Moreira and Mooney believed that the necrotrophic pathogensmay provide a significant biotic factor which indirectly modulates theoutcome of interspecific interactions at higher trophic levels [12-14].
The parasitoid A. colemani belongs to the sub-family Aphidiidae(Hymenoptera: Braconidae) which is a koinobiont [15-17]. Saljoqireported that the rate of parasitism by natural enemies was related tothe availability of the host, plant age, temperature, weather conditionsand cultural practices [18]. Saljoqi and van Emden reported thatM. persicae can attain very high densities on young plant tissues,causes water stress, wilting and reduced growth rate of the plant [19]. However, natural enemies can be used to limit survival of thehost. The female parasitoids choose hosts that provide the bestresources for their offspring, were they select hosts, and control thesex of their offspring by influencing fertilization [20-24]. The femalehymenopteran parasitoids regulate the release of sperm from thespermatheca while eggs pass through her genital tract; unfertilizedeggs develop into males while fertilized eggs develop into females[23,25,17]. Because fitness and size are strongly correlated withsize in female parasitoids, but not for males, female parasitoids canpreferentially oviposit male eggs in poorer quality host when providedwith a choice [26,22,27,17]. This research describes an experimentaimed at investigating whether the presence of the systemicpathogen, B. cinerea, on lettuce plant Lactuca sativa L. (Asteraceae:Compositae) influences the parasitization of an insect herbivore (thepeach potato aphid Myzus persicae) by the parasitoid A. colemaniwhich may provide a clue on the biocontrol of insect pest aphidM. persicae. We tested three hypotheses. First, that Myzus persicaereared on uninfected lettuce plants would be larger. Second, that A.colemani which emerges from an aphid host feeding on uninfectedplants would in turn be larger. Third, that decision to oviposit or notwould be influence by the presence or absence of B. cinerea infection.
Material and Methods
Experimental plants
Lettuce seeds (Tom Thumb), infected (obtained by inoculatinglettuce plants at the flower stage) and uninfected by B. cinerea, wereused for the experiment. Lettuce seeds were individually sown in90 (45 infected, 45 uninfected) 15cm pots filled with a vermiculitegrowing medium in a controlled environment (18-20°C, L: D 12-12,ambient humidity).
Infestation with Myzus persicae
Three weeks after germination all the plants were individuallyinfested each with 10 nymphs M. persicae. Myzus persicae were placedon the reverse side of the leaves using a small moist brush. Plants werethen covered with a transparent plastic dome for 48 h. Aphid colonieswere covered at all times by a vented plastic container, preventingescape of aphids or parasitoids.
Parasitization rate
The source parasitoid Aphidius colemani is a commerciallyavailable biocontrol agent, and was obtained from Koppert UK.Female A. colemani oviposit a single egg into aphid nymphs. Thelarva develops inside the body of aphid host until it kills the host.The experimental A. colemani were fed with honey and maintainedin vials at 5 °C in the laboratory. Prior to use parasitoids were allowedto acclimatize to room temperature for several hours. Immediatelyafter the adult aphids were removed from the plants, the nymphswere exposed to attack by five female A. colemani. Thirty infected anduninfected plants were attacked with the aphid natural enemy whilethe aphids on the remaining fifteen infected and uninfected plantswere allowed to serve as unattacked controls.
Rate of parasitoid attack
Emergence of mummies was determined by visual observationof the plastic doom. One week after parasitoid attack all the adult aphids were counted and collected into 200ml Eppendorf tubes andplaced on a laboratory bench for one week to allow the emergence ofparasitoids.
Size of M. persicae and A. colemani
Ten adult Myzus persicae were randomly selected from eachplant while all the resulting parasitoids from the mummies werecollected for the determination of hind tibia length. Both Myzuspersicae and Aphidius colemani were individually placed in a dropof 100% ethanol on a glass slide covered with a cover slip and hindtibia length measured using a micrometric eye piece attached to aNikon Olympus microscope (New York Microscope Company) at 50x magnifications.
Statistical analysis
All the experimental data were analysed using R [28]. Because thedata met assumptions of normality hind tibia length data were nottransformed prior to analysis. Attack rate and data counted from sexratio were analysed using a generalised linear model using binomialerrors [28].
Results
Size of M. persicae
Myzus persicae grown on plants uninfected by B. cinerea werelarger than those reared on infected plants after parasitoid attack onboth plants (Figure 1;F1,38 = 260.02, P < 0.001). In the control plantswhere aphids were not attacked by the parasitoids, aphids reared onuninfected plant were also larger than those reared on infected plants(F1,38 = 22.40, P <0.001).
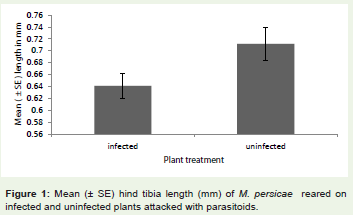
Figure 1: Mean (± SE) hind tibia length (mm) of M. persicae reared on infected and uninfected plants attacked with parasitoids.
Aphidius colemani size
The hind tibia length of parasitoid that emerged from aphidsreared on infected plants was significantly smaller than parasitoidgrown on aphids feeding on uninfected plants (F1, 59 = 134.10, P < 0.001). Female Aphidius colemani were significantly longer than maleAphidius colemani when reared on both infected (Figure 2; F1,59 =131.34, P < 0.001) and uninfected plants.
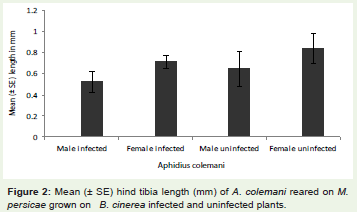
Figure 2: Mean (± SE) hind tibia length (mm) of A. colemani reared on M. persicae grown on B. cinerea infected and uninfected plants.
Sex ratio and parasitization rate by Aphidius colemani
The attack rate suffered by M. persicae was higher in uninfectedplants and also more A. colemani was recorded than on infectedplants (Figure 3; F1,39 = 4.20, P = 0.01). However, there was nosignificant effect of plant infection status on the sex ratio of emergingA. colemani, which has same proportion.
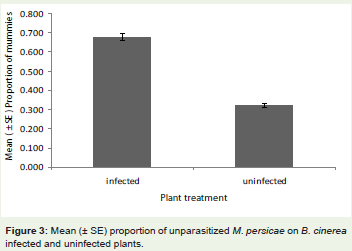
Figure 3: Mean (± SE) proportion of unparasitized M. persicae on B. cinerea infected and uninfected plants.
Discussion
The systemic pathogen appears to influence the dynamics ofthe insect herbivore and its natural enemy. More M. persicae andparasitoids Aphidius colemani were grown on lettuce plants freefrom infection by B. cinerea. The aphids and emerging parasitoidswere significantly larger when reared on uninfected plants. Similarparasitoid sex ratio, 50:50 was found between parasitoids emergingfrom both infected and uninfected plants. The study found thatsize of the aphid is a measure of host quality, where host quality isa linear function of size and determines the ability of the developingparasitoid to obtain the required nutritional resources from the host this was similarly, reported by Sequeira and Mackauer [29] and Odeet al. [22]. The results showed that female A. colemani oviposit morein hosts reared on uninfected plants, probably because their offspringhave a higher probability of survival this was similarly reported byRoitberg et al. and Ode et al. 2005 [30,22]. Larger hosts have morefood and provide a better resource for the growth and developmentof offspring, which in turn influences the survival and fecundity ofthe adult parasitoid.
The study showed that a high number of parasitoid mummies were recorded in aphids reared on B. cinerea free plants, which isan indication that presence of B. cinerea exert a negative influenceon the interactions between M. persicae and their natural enemyA. colemani. Previous studies showed that this negative interactionoccur because Botrytis cinerea can lowered the fitness of the aphidsmaking it a poor-quality hosts resulting in a high mortality rate ofthe parasitoid [31]. This occurs when the B. cinerea induce the plantto synthesize secondary metabolites which may have toxic effects onthe aphids. Therefore, the foraging parasitoid discriminate betweenfeeding on aphid reared on infected and uninfected plants whetherto feed, oviposit or to ignore, because host choice correlates withoffspring performance which has a direct impact on the parasitoidhost choice [32,31,27]. However, in the present study, parasitoidswere not allowed a choice between infected and unifected hosts, andas a result it is not possible to state categorically if host preference islikely to have influence the differences in attack rate.
In agreement with the present experiment the work of Jarosik etal. [33], Harri et al. [34]; Rehman and Powell [24] have shown thatA. colemani emerging from small aphids, showed a significant malebiased offspring sex ratio. The shift in emergence sex ratio may bethe results of a significantly higher mortality of female progeny insmaller hosts. Female A. colemani often have a more strict nutritionalrequirements compared to males and may therefore reach a largersize than males even in a hosts of equal size, where the female haveextended period of larval development on growing hosts which enablesthem to consume more resources than the males [29,35,15,36,37]. Inthe present study more aphids and parasitoids with larger size whererecorded from uninfected plants. As reported by Charnov et al. [20]and Paul et al. [38], higher female mortality in small aphids althoughnot measured in the present study normally occurs due to a shortageof resources during the early stages of female development. The sexratio determined by female choice at oviposition may as a result bealtered by differential mortality of the sexes. Therefore a biased sexratio may result either from the female’s control of fertilization atoviposition, or may result from the differential mortality of male andfemale offspring during early stages of development [20,38].
Overall the result of the present study has confirmed theinfluence of B. cinerea on the parasitization M. persicae by parasitoidsA.colemani. The observations of the experiment provide furtherevidence that healthy plants free from B. cinerea infection providebetter quality food resources for the aphid host which encourage fitnessand results in larger sized offspring. The study found that infectedplants which were infested with aphids showed reduced aphid andparasitoid population counts which indicated the negative effect of B.cinerea on the population dynamics of both herbivore and its naturalenemy. Therefore the negative effect of B. cinerea on the interactionbetween aphid and its natural enemy parasitoid determined in thepresent study may be very valuable in understanding interaction athigher trophic level and may also provide a clue for design of diseasemanagement strategies.
Acknowledgement
Special thanks to Sue Ramsey and Lee W. Y. for their technicalassistance. And Amanda Livermore for providing me with aphidsand guidance on aphid rearing.
References
- Blackman RL, Eastop VF (2000) Aphids on the world’s Herbaceous Plants and Shrubs, Wiley, Chichester, England.
- Eisen J (2010) Genome sequence of the pea aphid Acrythosiphon pisum. Plos Biol 8: e 1000313.
- Pell JK, Pluke R, Clark SJ, Kenward MG, Anderson PG (1997) Interactions between two aphid natural enemies, the entomopathogenic fungus Erynia neoaphidis Remaudiere and Hennebert (Zygomycete: Entomophothorales) and the predatory beetle Coccinella septempunctata. L. (Coleoptera: Coccinellidae). Invertebr Pathol 69: 261-268.
- Mouttet R, Bearez P, Thomas C, Desneux N (2011) Phytophagous arthropods and a pathogen sharing a host plant: Evidence for Indirect Plant-Mediated Interactions. Plos One 6: e18840.
- Hatcher PE, Paul ND, Ayres PG, Whittaker JB (1995) Interactions between Rumex spp. herbivores and a rust fungus: the effects of Uromyces rumicis infection on leaf nutritional quality. Functional Ecology 9: 97-105.
- Gwynm DM, Callagham A, Gorham J, Walters KFA, Fellowes MDE (2005) Resistance is costly: trade-offs between immunity, fecundity and survival in the pea aphid. Proc Biol Sci 272: 1803-1808.
- Hazell SP, Fellowes MDE (2009) Intra-specific variation affects the structure of the natural enemy assemblage attacking pea aphid colonies. Ecological Entomology 34: 34-42.
- Trumble JT (1982) Within plant distribution and sampling of aphids (Homoptera: Aphididae). On Broccoli in southern California. Journal of Economic Entomology 75: 587-592.
- Fellowes MDE, Van Alphen JJM, Jervis M (2005) Foraging behaviour. In Mark AJ (ed.) Insects as Natural Enemies: A Practical Perspective. Pp 1-71. Springer, Dordrecht, the Netherlands.
- Yahaya SM, Fellowes MDE (2013) Host plant infection by Botrytis cinerea affects the interaction between aphids and their parasitoids. Journal of Biological Science and Bioconservation 5: 111-123.
- Hatano E, Kunert G, Michaud JP, Wolfgang W, Weisser W (2008) Chemical cues mediating aphid location by natural enemies. Eur J Entomol 105: 797-806.
- Persson L (1999) Trophic cascades: abiding heterogeinity and the trophic level concept at the end of the road. Oikos 85: 385-397.
- Aquilino KM, Cardinale BJ, Ives AR (2005) Reciprocal effects of host plant and natural enemy diversity on herbivore suppression: an empirical study of a model tritrophic system. Oikos 108: 275-282.
- Moreira X, Mooney KA (2013) Influence of plant genetic diversity on interactions between higher trophic levels. Biol Lett 9: 20130133.
- Godfray HCJ (1994) Parasitoids: Behavioral and Evolutionary Ecology. Princeton University Press, Chichester, UK.
- Colinet H, Salin C, Boivin G, Hance TH (2005) Host age and fitness-related traits in a koinobiont aphid parasitoid. Ecological Entomology 30: 473-479.
- Kant R, Minor MA, Trecwick SA (2012) Fitness gain in a koinobiont parasitoid Diaretiella Rapae (Hymenoptera: Braconidae) by parasitizing hosts of different ages. Journal of Asia -Pacific Entomology 15: 83-87.
- Saljoqi AR (2009) Population dynamics of Myzus persicae(Sulzer) and its associated Natural enemies in spring potato crop, Peshawar Pakistan. Sarhad Journal of Agriculture 25: 564-571.
- Saljoqi AUR, Van Emden HF (2003) Selective toxicity of different granular insecticides to the peach-potato aphid, M. persicae (Sulzer) (Homoptera: Aphididae) and its parasitoid, A. matricariae Haliday (Hymenoptera: Aphidiidae) in two differentially resistant potato cultivars. Journal of Biological Sciences 3: 215-227.
- Charnov EL, Los-den Hartogh RL, Jones WT, Van den Assem J (1981) Sex ratio evolution in a variable environment. Nature 289: 27-33.
- King BH, Seidl SE (1993) Sex ratio response of the parasitoid wasp Muscidifurax raptor to other females. Oecologia 94: 428-433.
- Ode PJ, Hopper KR, Coll M (2005) Oviposition vs offspring fitness in Aphidius colemani parasitizing different aphid species. Entomologia Experimentalis et Applicata 115: 303-310.
- He XZ, Wang Q (2006) Host age preference in Aphidius ervi (Hymenoptera: Aphididae). Journal of New Zealand Plant Protection Society 59: 195-201.
- Rehman A, Powell G (2010) Host selection behaviour of aphid parasitoids (Aphidiidae: Hymenoptera). Journal of Plant Breeding and Crop Science 2: 299-311.
- Rabelling C, Lino-Neto J, Cappellari SC, Dos-Santos IA, Mueller UB, et al. (2009) Thelytokous parthenogenesis in fungus-gardening Ant Mycocepurus smithii (Hymenoptera: Formicidae) Plos One 4: e6781.
- Morris R, Fellowes M (2002) Learning and natal host influence host preference, handling time and sex allocation behaviour in a pupal parasitoid. Behavioural Ecology and Sociobiology 51: 386-393.
- Bilu E, Hopper KR, Coll M (2006) Host choice by Aphidius colemani: effects of plants, plant-aphid combinations and the presence of intra-guild predators. Ecological Entomology 31: 331-336.
- Crawley MJ (2005) An Introduction Using R. Wiley. United Kingdom.
- Sequeira R, Mackauer M (1992b) Covariance of adult size and development time in the parasitoid wasp Aphidius ervi in relation to the size of its host Acrythosiphon pisum. Evolutionary Ecology 6: 34-44.
- Roittberg BD, Boivin G, Vet LEM (2001) Fitness, parasitoids and biological control: an opinion. Can Entomol 133: 429-438.
- Kruess A (2002) Indirect interaction between a fungal plant pathogen and a herbivorous beetle of the weed Cirsium arvense. Oecologia 130: 563-569.
- Guevara R, Rayner ADM, Reynolds SE (2000) Orientation of specialist and generalist fungivorous ciid beetles to host and non-host odours. Physiological Entomology 25: 288-295.
- Jarosik V, Holy I, Lapchin L, Havelka J (2003) Sex ratio in the aphid parasitoid Aphidius colemani (Hymenoptera: Braconidae) in relation to host size. Bull Entomol Res 93: 255-258.
- Harri SA, Kraus J, Muller CB (2008) Fungal endosymbionts of plants reduce lifespan of an aphid secondary parasitoid and influence host selection. Proc Biol Sci 275: 2627-2632.
- Sequeira R, Mackauer M (1993a) Nutritional ecology of an insect host-parasitoid association: the pea-aphid Aphidius ervi system. Ecologist 73: 183-189.
- Mackauer M (1996) Sexual size dimorphism in solitary parasitoid wasps: influence of host quality. Oikos 76: 265-272.
- Lykouressis D, Garantonakis N, Perdikis D, Fantinou A, Mauromoustakos A (2009) Effect of female size on host selection by a koinobiont insect parasitoid (Hymenoptera: Braconidae: Aphidiinae). Eur J Entomol 106: 363-367.
- Ode P, Berenbaum MR, Zangeri AR, Hardy ICW (2004). Host plant, host plant chemistry and the polembryonic parasitoid Copidosoma sosares: indirect effects in a tritropic interaction. Oikos 104: 388-400.