Phylogenetic Relationships of Host Plants of Whitefly, Bemisia tabaci (Genn.)
Y.S. Rathore1 and S.N. Tiwari2*
Corresponding author: S.N. Tiwari, Department of Entomology, G.B. Pant University of Agriculture & Technology, Pantnagar-263145, India,; E-mail: drsntiwari@gmail.com
1Indian Institute of Pulses Research, Kanpur - 208024 (UP), MIG 110, Green Belt, Indira Nagar, Kanpur 208026 (UP)
2Department of Entomology, G.B. Pant University of Agriculture & Technology, Pantnagar-263145, Uttarakhand
Citation: Rathore YS, Tiwari SN. Phylogenetic Relationships of Host Plants of Whitefly, Bemisia tabaci (Genn.). J Plant Sci Res. 2014;1(3): 111.
Copyright © 2014 Y.S. Rathore et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Plant Science & Research | ISSN: 2349-2805 | Volume: 1, Issue: 3
Submission: 04/08/2014; Accepted: 11/10/2014; Published: 14/10/2014
Abstract
Bemisia tabaci (Gennadius) is capable of feeding on 910 plant species, belonging to 114 families and 71 orders globally. It prefers more dicotyledonous plants than the monocot ones. Fifty percent plants fall in Lignosae group (fundamentally woody plants) and 42.31percent in the Herbaceae group (fundamentally herbaceous plants) of dicotyledons. Monocotyledons shared 7.25 percent from 17 families and 12 orders. Six phylogenetic lineages showed prominence in dicotyledons and two in monocotyledons. Since B. tabaci has such a large number of hosts, ranging through many families and orders makes it undoubtedly a polyphagous pest and truly said world’s mostdangerous insect pest.
Keywords: Bemisia tabaci; Lignosae; Herbaceae; Host plant; Host range; Phylogeny; Phylogenetic relationship; Whitefly
Introduction
Bemisia tabaci (Gennadius) (Homoptera: Aleyrodidae) is commonly known as tobacco whitefly, sweet potato whitefly, cotton whitefly, etc. It has been reported from almost all the continents except Antarctica [1] (issg database). Over 900 host plants have been recorded and it transmits as many as 111 virus species and thus has been nominated as world’s worst invader (issg database). B. tabaci is a phloem feeder and weakens plants by desapping cell sap or photosynthates and lowering their vitality. Symptoms of its attack vary with the host species. In pulse crops, the disorder wasreferred to as downward cupping of leaves [2], in squash, silver leaf [3], uneven ripening of tomato [4] and in cole crops as stem streaking [5]. Secondary damage is caused through honeydew excretion which promotes sooty mold development blocking sunlight and thus reducing photosynthesis.
Feeding also causes increased stomatal resistance (impaired gas exchange), reduces transpiration and photosynthesis rate and chlorophyll content in tomato leaves [6]. Reduction in photosynthesis rates in cotton causes accumulation of soluble sugar, suggesting interference in the export of fixed carbon [6]. Plants accumulate pathogenesis-related (PR) proteins in response to B. tabaci feeding [7,8]. Infested tomato produced higher levels of PR proteins, B-1, 3-glucanase, chitinase, peroxidase PR2 and PR4 both locally and systematically [9,10]. Source-sink manipulations were demonstratedon cantaloupe [11] where B. tabaci aggregation altered free amino acid composition and elevated their total concentration in the phloem sap. B. tabaci, therefore, influences physiology, biochemistry and anatomy of the host plants [6]. Viewing its vast distribution and acceptability of large number of host species, it was thought pertinent to analyse its feeding preferences particularly related with plant phylogeny.
Materials and Methods
Nine hundred plant species reported in the literature as a host of B. tabaci were considered for discussion on the phylogenetic relationship. Additional plants on which the insect was intercepted at the plantquarantine were also included. Therefore, the total number of plants involved in the present study was 910. However, host races, biotypes and populations of B. tabaci and their geographical distribution were kept aside because of paucity of information on all the aforesaid aspects. Hering [12] proposed classification on phylogeographical basis and coined the terms “xenophobic” and xenophilic” which refer to rejection and acceptance of plants indigenous to a region, respectively. Thorteinson [13] hold opinion that some insects devour indiscriminately while others are particularly associated with specific species of plant or each insect species is associated with a group of plants which were designated as the food plant range. Food plants of the insect, therefore, are certainly correlated with natural taxonomic plant groupings, although in some insects it may happen without special regard to botanical affinity.
To study the phylogenetic relationships of host plants of B. tabaci, the classification of Hutchinson [14] was adopted which divided phylum Angiosperm in subphylum dicotyledons and monocotyledons. Dicotyledons are grouped in Lignosae (fundamentally woody group) and Herbaceae (fundamentally herbaceous group). Food plant of B. tabaci were aligned to the respective taxonomic group (orders) and presented on evolutionary basis.
Results and Discussion
In the literature, 910 plant species has been reported as host of whitefly. Out of these, 844 are dicotyledons while 66 belong to monocotyledons. These facts clearly indicate that dicotyledons are more preferred by B. tabaci as compared to monocotyledons. Among dicotyledons, this insect preferred more plants belonging to Lignosae (459 or 50.43%) - as compared to Herbaceae (358 or 42.31%). These results are not in agreement with De Barro [15] who mentioned that B. tabaci prefers short lived herbaceous hosts, including numerous agricultural and horticultural species of dicotyledons. Monocotyledonous plants had only 7.25% share of the total number of plants. The percentage of families and orders in various groups also followed the same trend (Table 1).
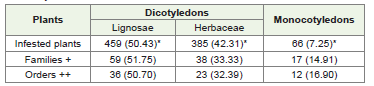
Table 1: Number of B. tabaci infested plants observed in dicotyledons andmonocotyledons.
Among dicotyledons, plants were preferred from most primitive to most advanced orders in both Lignosae and Herbaceae. Magnoliales, the most primitive in Lignosae (Liliodendron tulifera) and Ranales the most primitive in Herbaceae (Clematis linguisticifolia, Clematis spp., Ranunculus longstdorfii, R. muricatus, Ranunculus spp.) contain 1 and 5 plants, respectively. Magnoliales and Ranales evolved from common but remote ancestor [14]. Zomlefer [16] put 6 families, namely, Illiciaceae, Magnoliaceae, Annonaceae, Aristolochiaceae,Lauraceae and Piperaceae in the order Magnoliales. If we accept the classification for a while, then 17 plant species harboured by B. tabaci. However, Hutchinson [14] kept Piperaceae in Herbaceae and elevated other families as orders. He also described 54 orders in the group Lignosae. Out of these, plants of 59 families were infested which belonged to 36 orders. In other words, one half of the plants were preferred by B. tabaci which were primarily and predominantly woody plants. Dilleniales is remotely related to Magnoliales and is showing a connecting link between Rosales, Bixales, Theales and Guttiferales. However, Dilleniales were not at all infested by B. tabaci. On the other hand, Rosales, Bixales and Guttiferales had good number of infested plants. Rosales is placed at 6th on evolutionary scale and it included 13 plants in the family Rosaceae and one in Chrysobalanceae. Later family has now been merged in Rosaceae. Rosaceae is generally accepted as monophyletic group where saponins and cyanogenic glucosides are commonly present [16]. Plants reported as hosts are Chrysobalanus orbicularis (coco plant), Fragaria vesca, Heteromeles solicifolia, Malus domestica (apple), Potentilla spp., Pyrus calleryana (pear), Pyrus communis, Pyrus mamorensis, Pyrus spp., Rosa chinensis, Rosa gallica, Rosa spp., Rosa centifolia and Rubus fruticosus (blackberry) (Table 2, Figure 1).
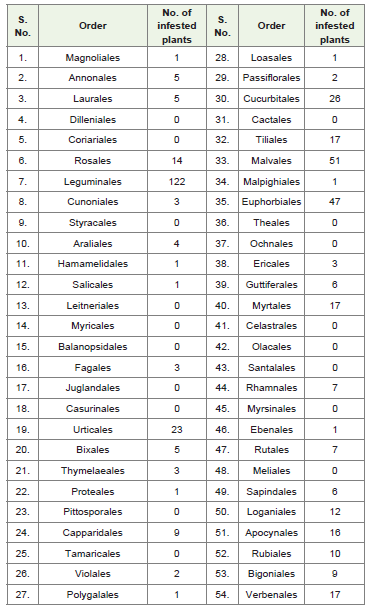
Table 2: Number of infested plants observed in various orders of Lignosaegroup of dicotyledons (from primitive to advanced orders).
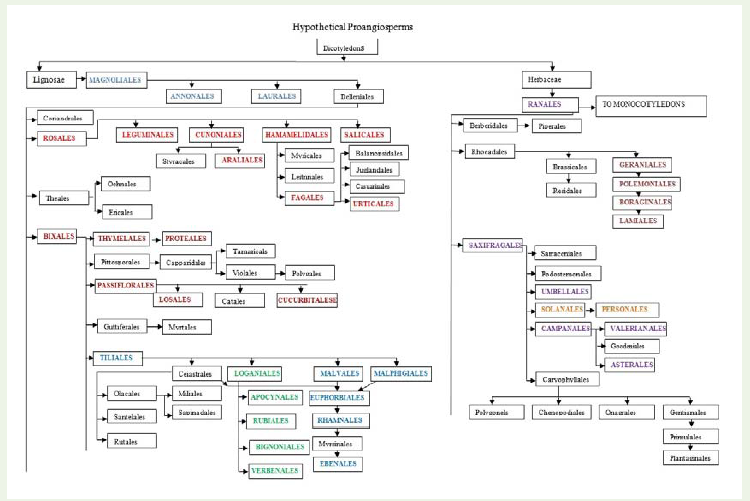
Figure 1: Diagram showing the probable phylogeny and relationships of the order of dicotyledones (adapted from Hutchinson, 1973). Host orders have been super imposed by bold capital font to relate the phylogenetic relationship.
Next large order Leguminales is placed at 7th on evolutionary scale and hosted 122 plants (Table 2). This group of plants was derived from Rosales stock and represented by three important families, namely, Caesalpiniaceae, Mimosaceae and Fabaceae (Papilionaceae) embracing 18, 8 and 96 plants, respectively. Fabaceae contains many important grain legume crops viz., peas, broad bean, groundnut, soybean, lentils, clover, chickpea, pigeonpea, green gram and black gram. Fabaceae is characterized by presence of various alkaloids and cyanogenic glucosides. Tissues also possess calcium oxalate crystals.
Rosales stock further gave two distinct lineages, of which one leads to Cunoniales-Styracales and Araliales (Figure 1). The other lineage gave rise to Hamamelidales which is closely allied to Rosales. Among Cunoniales 3 plants viz., Ribes spp., R. cynosbali and R. uva-crispa hosted by B. tabaci. Styracales had no host but Araliales provided 4 host plants from two families viz., Araliaceae (Hedra helix) and Caprifoliaceae (Loniara spp., L. etrusca, L. japonica). Salicales was represented by one host (Salix spp.) of family Salicaceae. Sevenorders evolved from Hamamelidales (Figure 1). Out of these, one phylogenetic lineage was Rosales-Hamamelidales-Fagales-Urticales. Last three orders possessed 1, 3 and 23 host species, respectively(Table 2). Four families viz., Cannabaceae, Moraceae, Ulmaceae and Urticaceae fall under the order Urticales. Thorne [17] opined that the monophyletic Urticaceae s.s. is derived within the ‘Moraceae’ and the variously circumscribed ‘Moraceae’ and ‘Cercopiaceae’ as probablyparaphyletic. Author, therefore, suggested the inclusion of Moraceae and Cercopiaceae within the expanded Urticaceae s.l.
Order Bixales occupied the 20th position on evolutionary scale and whitefly was found on 5 plant species (Table 2, Figure 1) belonging to three families, namely, Bixaceae (Cochiospermum planchoni), Cistaceae (Cistus spp., C. salvifolius, Helianthemum spp.)and Flacourticeae (Raosonia lucida). Orders Tiliales, Passiflorales and Thymelaeales derived from Bixales and in turn Cucurbitales and Losales evolved from Passiflorales and Proteales from Thymelaeales (Figure 1). Guttiferales has offshoot evolution before Tiliales and as a further step ended in Myrtales. Tiliales, Passiflorales and Thymeleales got 17, 2 and 3 plant species, respectively. Tiliales got host plants in three families viz., Bombacaceae (2), Sterculiaceae (6) and Tiliaceae (9). Zomlefer (1994) put all these families in order Malvales. Judd et al. [18] suggested that Bombacaceae is likely paraphyletic and probably should be merged with the Malvaceae. Sterculiaceae is closely related to the Tiliaceae, Malvaceae and Bombacaceae; the circumscriptions of these families in relation to one another is problematic. Sterculiaceae is known for the presence of tannins, sterculic and malic acids and various alkaloids e.g. caffeine, theobromine [16]. Cucurbitales and Losales had 26 and 1 plant species, respectively. Twenty four plants fall in the family Cucurbitaceae and one each in Begoniaceae and Caricaceae. According to Thorne [17] family Cucurbitaceae is generally allied with Begoniaceae. Cucurbitacin (tetracyclic triterpenoids), bitter tasting substances and various alkaloids are typically present in this family. One plant species was observedin Losales in family Turnaraceae and one in Proteales in family Proteaceae (Table 2, Figure 1).
The other adopted phylogeny of B. tabaci was Bixales-Tiliales-Malvales-Euphorbiales-Rhamnales-Ebenales. Tiliales is fairly advanced group. Malvales is a natural and climax group with a single family Malvaceae. Sterculic and malvalic acids are commonly present in the members of this family. Euphorbiales is a heterogenous groupand has a single family Euphorbiaceae which probably has been derived from several stocks [14]. Phylogenetic relationship, therefore, has been much disputed. Range of morphological variation is so great that many authors hypothesize a paraphyletic origin of the family. Various alkaloids, cyanogenic glucosides and tannins are commonly present in the family. Tissues are with oxalate crystals [16]. Rhamnales are closely related to Celastrales which has no host of B. tabaci. Ebenales came up from Rhamnales via Myrisnales which was not infested (Figure 1). Malvales and Euphorbiales together contributed as many as 98 host species and Malpighiales, Rhamnales and Ebenales had 1, 7 and 1 plant species, respectively (Table 2).
The other small lineage was Capparidales-Violales-Plygalales which had 9, 2 and 1 plant species, respectively. The other short lineagewas Rutales-Sapindales. Seven plant species in Rutales belonged to Rutaceae and 6 plants in Sapindales fall in three families, namely, Aceraceae (2), Anacardiaceae (3) and Sapindaceae (1). Rutaceae is characterized by having aromatic parts (due to ethereal oils). In triterpenes various alkaloids are also present like calcium oxalatecrystals. The Rutaceae are allied with families in the Rutales-Sapindale complex such as Sapindaceae, Meliaceae and Anacardiaceae. The last phylogeny was observed among the advanced climax groups. Loganiales gave rise to Rubiales, Apocynales, Verbenales and Bignoniales. They had 12, 10, 16, 17 and 9 host species, respectively,totaling to 64. Family structure was Loganiaceae (1) and Oleaceae (11) in Loganiales; Rubiaceae (10) in Rubiales; Apocynaceae (10) and Asclepiadaceae (6) in Apocynales; Verbenaceae (17) in Verbinales; and Pedaliaceae (4) and Bigoniaceae (5) in Bignoniales (Table 2). Since B. tabaci feeds on 459 plant species only in Lignosae, its hostsare distributed in many orders. However, clear phylogenetic lines appear as follows:
1. Rosales – Leguminales which further expanded to 6 other orders,
2. Bixale – Passiflorales, Thymeleales which gave rise to Cucurbitales, Losales and Proteales,
3. Bixale – Tiliales-Malvales-Euphorbiales-Rhamnales-Ebenales, and
4. Loganiales-Rubiales, Apocynales, Verbenales, Bignoniales.
The other orders which showed small phylogenetic sequences seem to have isolated infestation. It is difficult to explain why their progenitors had no infestation while, the orders evolved from suchprogenitors had abundant number of plants susceptible to B. tabaci. It could be due to different geographical distribution and unconducive environment for development and survival of whitefly. However, B. tabaci was reported to feed on plants from primitive orders to the most advanced orders in Lignosae group of dicotyledons. In theevolutionary system, the order Ranales took first place as being the most primitive of the fundamentally herbaceous group. Relationship of whitefly to herbaceous group of plants was bit different than the Lignosae. Here plants occupied 32.39% of orders and 33.33% families as compared to 50.70 and 51.75% in Lignosae, respectively. Among the Herbaceae, 85.90% plants were contributed by the orders Chenopodiales, Umbellales, Asterales, Solanales, Personales, Geraniales, Lamiales and Brassicales. Except 5, all other orders had representatives. However, some of the phylogenetic lineages were more prominently represented. For example, Ranales-Saxifragales-Solanales-Presonales had 32.80% infested plants followed by Ranales-Saxifragales-Camponales-Asterales and Valerianales which shared 29.90% plants. Phylogenetic lineage from Ranales to Lamiales and Ranales to Chenopodiales got more that 16% of plant species.However, the order Brassicales which is an advanced group had 14 plants of the family Brassicaceae (Cruciferae) while, its progenitor Rhodeales showed no infestation (Table 3, Figure 1).
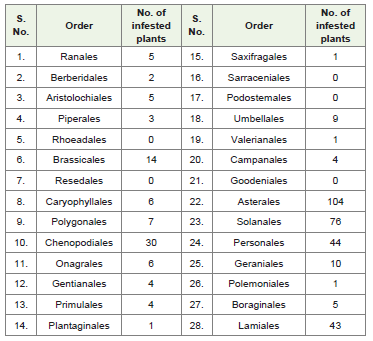
Table 3: Number of infested plants observed in various orders of Herbaceaegroup of dicotyledons (from primitive to advanced orders).
Phylogenetic tree of monocotyledons as described by Hutchinson indicates that representatives of the orders of Butanales and Alismatales are closely allied to the family Ranunculaceae of the order Ranalesand considered this group as the monophyletic. Ranunculaceae possesses perennial and annual herbs or shrubs or climbers. On the other hand, plants belonging to Alismataceae are perennial or annual marsh or aquatic herbs or rarely with floating leaves. Alismataceaeplants resemble to Ranunculaceae. There is some divergence of opinion whether the resemblance is due to real relationship or merely to parallel development in the two great divisions. The Alismataceae was cited as primitive group of extant monocots [16]. Lotsoy [19] believed that monocotyledons have diphyletic origin, i.e., some families in dicotyledons and the remainder of monocotyledons from the hypothetical Proranales. Alismatales gave rise to Commelinales which further developed in Liliales and Bromeliales (Table 4 and Figure 2). Heterenthera spp. is a host of B. tabaci and belongs to family Pontedriaceae. Hutchinson [14] placed this family in Liliaceae. However, Dahalgren et al. [20] and Thorne [17] regarded considerable variation in the phylogenetic scheme of this family and allied withdiverse groups as the Bromeliaceae. Zomlefer [16] placed this family in Bromeliales. If we accept it, then there is one phylogenetic lineage Alismatales-Commelinales-Bromeliales-Zingiberales. Except Liliales, no other order derived from Commelinales or Alismatales has any representative as host of B. tabaci. Liliales is a large group from which prolific evolution has taken place. Sixteen orders have originated from Liliales and out of them 8 have shown the host plantsof B. tabaci. Order like Orchidales has only one plant. Phylogenetic lineage is developing from Liliales-Agavales-Palmales which had 10 host plants. Arales and Dioscoreales are also derived directlyfrom Liliales. Graminales came up via Juncales. The three orders, namely, Arales, Dioscoreales and Graminales constituted 54.5% of the total number of infested plants. Family Araceae (Arales) and Dioscreaceae (Dioscoreales) are derived from Liliaceae while, Poaceae or Gramineae (Graminales) perhaps developed parallelwith Cyperaceae through Juncaceae stock from the Liliales. Order Amaryllidales also possesses only one family Amaryllidaceae which is very close to Liliaceae, and two are differentiated on the basis of superior or inferior ovaries. Iridales also have one family Iridiaceaeevolved separately from Liliaceae and on separate line [14]. Botanists generally concur that Iridaceae are related to the Liliaceae s.s. and allied families of the Liliales [16]. Amaryllidales and Iridales have one and two host plants, respectively. Alismatales-Commalinales-Bromeliales and Zingiberales phylogenetic lineage contributed 25.8% plants of the monocotyledons. Therefore, two phylogenetic lineages are prominent. In some of the orders like Orchidales which are comparatively advanced ones, infestation appears to be isolated as in between them links are not available (Table 4, Figure 2).
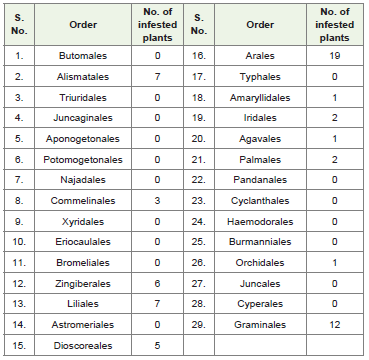
Table 4: Number of infested plants observed in monocotyledons (from primitiveto advanced orders).
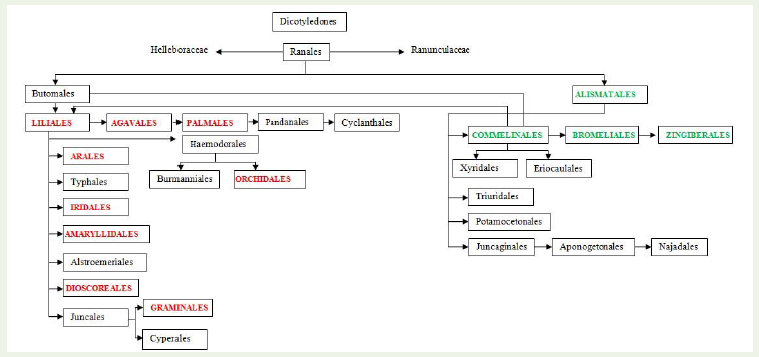
Figure 2: Diagram showing the probable phylogeny and relationships of the order of Monocotyledons (Adapted from Hutchinson, 1973). Host orders have been superimposed by bold capital font to relate phylogenetic relationship.
If we consider 3-4 top orders in three groups, namely, Lignosae, Herbaceae and monocotyledons, it was found that Leguminales (122) topped in Lignosae followed by Malvales (51), Euphorbiales (47) and Cucurbitales (26) and so on. Leguminales is a prolific group derived from Rosales (14) stock, through Caesalpiniaceae (18), Mimosaceae(8) and Fabaceae (96). Caesalpiniaceae is closely related to primitive family Rosaceae, the Mimosaceae to be relatively more advanced, and the Fabaceae to be the climax group, providing most plants of economicimportance (e.g. peas, beans, lentil, clover, rosewood, chestnut, etc.). Leguminous plants are famous for fixing nitrogen in the soil. Malvales has single family Malvaceae which is characterized by herbaceous to softly woody stems, often fibrous. Its useful plants are Hibiscus, Gossypium, Abutilon, etc. According to Zomlefer [16]order Malvales consists of 4 families, namely, Malvaceae, Bombacaceae, Sterculiaceae and Tiliaceae. Judd et al. [18] suggested that Bombacaceae is likely paraphyletic and probably should be merged with Malvaceae. Euphorbiales contains single family, i.e., Euphorbiaceae. A composite (heterogeneous) family probably derived from severalstocks such as Bixales, Tiliales, Celastrales and perhaps Sapindales [14]. Phylogenetic relationships have been much disputed. Other families (such as Boxaceae) sometimes have been included. Morpho logical variation is so great that many authors hypothesize a polyphyleticorigin of the family [16]. It contains many economic plants. Cucurbitales are undoubtedly related to Passifloraceae. Hutchinson classified 4 families in Cucurbitales and out of these B. tabaci infested 3, namely, Cucurbitaceae (24), Begoniaceae (1) and Caricaceae (1). Cucurbitaceae encompasses useful plants like melon, cantaloupe, cucumber, squash, zucchini, bottle gourd, sponge gourd, bitter gourd, Mamordicha spp., and Trichosanthes spp., etc.
Analysis of plants in Herbaceae revealed that order Asterales (104) topped the group followed by Solanales (76), Personales (44), Lamiales (43). Chenopodiales also comprised of 30 host plants. Asterales mainly derived from Campanulaceous stock. It is known to have single family (Asteraceae) but one of the largest angiospermfamilies [16]. It has wide range of plants generally used as food, medicines, insecticides, etc. Out of 4 families of Solanales, two families viz., Solanaceae (55) and Convolvulaceae (21) contain host plants of B. tabaci. Many plants like potato, tomato, eggplant, chilies, jasmine, cherry, Datura, petunia, etc. belong to Solanaceae family and sweet potato, morning glory etc. to Convolvulaceae, respectively. Personales is a large natural order, largely herbaceous, comprising7 families, out of which B. tabaci has been found feeding on representatives of 3 families, namely, Acanthaceae (27), Gesneriaceae (2) and Scrophulariaceae (15). The phylogenetic relationship of Acanthaceae are also controvertial having affinities with groups likeScrophuliaceae, Gerinericeae, Verbenaceae and Lamiaceae [21,22]. Zomlefer [16] put Oleaceae (11), Scrophularaceae (15), Bignoniaceae (5), Plantaginaceae (1), Lentibularaceae (00), Acanthaceae (27), Verbenaceae (17) and Lauraceae (2) in one order Scrophulariales.Lamiales (43) possesses 4 families out of which B. tabaci feeds upon the plants of two families viz., Globulariaceae (1) and Lamiaceae (42). Family Lamiaceae is regarded highly advanced family of dicotyledons. It is noted for their secretion of oil used in perfumery.Useful products are mint, lavender oil, culinary herbs, artichoke, etc. Chenopodiales (30) is further reduction from Caryophyllaceous stock. This order is comprised of 11 families, however, B. tabaci infested representatives of 3 families only, namely, Chenopodiaceae (8), Amaranthaceae (21) and Phytolaccaceae (1). Plants like spinach,sugarbeet, ornamental plants and phytolacca belong to these families. The family Chenopodiaceae is generally considered to be closely allied with Amaranthaceae [23]. Rodman [24] observed that two taxa may be sister groups or the Amaranthaceae may be derived withinChenopodiaceae.
Among the monocotyledons, the order Arales (19) reported to have maximum number of plants followed by Graminales (12). The order Arales belongs to division Carolliferae and Graminales to Glumiflorae. Arales has two families but B. tabaci reported to infest plants belonging to the family Araceae, which is monophyletic inorigin i.e., from the stock of tribe Aspidistreae of Liliaceae. Acocasia, Amorphophalles, Antherium, Caladium, Colocasia, Philodendron, Syngonium, etc. comprised of this family. Order Graminales contributes one family (Poaceae or Graminae). This is one of the most important family which provides food as cereals and millets for human beings and forage for animals. Bemisia tabaci is reportedto feed upon Crox lacryma-jabi, Cynodon dactylon, Hemarithria compressa, Oplismenus burmanii, Oryza sativa, Pennisetum americanm, Pennisetum spp., Saccharum offcinarum, Sorghum bicolor, S. helepense, Urochloa adspersa and Zea mays.
B. tabaci has large number of host species across the large number of orders and families. Phylogenetic lineages are evident in all the groups. However, the distribution of host species, sequence of their natural occurrence and environmental influence on biology and development of B. tabaci in a given climate/nitch are not allowingclear cut presentation of different lines of insect-plant relationships. Keeping in view the classification of [13], it can be said that B. tabaci is a polyphagous insect and highly dangerous to crops.
Conclusions
1. Manifestation of host plants through diverse groups evidentially indicated that in some cases phylogenetic relationship is prominent than the others.
2. Distribution of host plants across the ecosystems or ecozones intermingled the information on seasonal abundance of B. tabaci and presence and suitability of host during active period of the pest.
3. Lack of information on type localities of plants, especially the forest plants and trees, which helps in sustaining the biodiversity not only in the forest ecosystem but also helps in stabilizing the agro-ecosystem.
4. To manage B. tabaci in agro-ecosystems one has to be vigilant for the presence of collateral and alternate hosts in nearby ecosystems also. Presence of meager population of B. tabaci in a nitch may help in population eruptions at favourable times.
5. Information on genetic diversity and ecological diversity will help in making phylogenetic chart which can predict that if you need to venture in new areas, the possibility is that B. tabaci may welcome you.
Acknowledgements
Authors are extremely grateful to Dr. Shanker Lal, former Economic Botanist (Legumes), U.P. Institute of Agril. Sciences, Kanpur and former Director of Indian Institute of Pulses Research, Kanpur for critically going through the manuscript and making necessary corrections.
References
- Issg Detabase: Ecology of Bemisia tabaci.
- Kooner BS, Cheema HK, Rubaljot K (2006) Insect pests and their management. In Advances in Mungbean and Urdbean. Eds. Masood Ali and Shiv Kumar 2006, IIPR: 335-340.
- Yokomi RK, Hoelmer KA, Osborne LS (1990) Relationships between the sweet potato whitefly and the squash silver leaf disorders. Phytopathlogy 80: 895-900.
- Schuster DJ, Muller TF, King JB, Price JF (1990) Relationship of the sweetpotato whitefly to a new tomato fruit disorder in Florida. Hort Science 25: 1618-1620.
- Brown JK, Lastra R, Bird J (1991) First documentation of whitefly transmitted Gemini viruses causing widespread disease in cotton, tobacco and tomato in Puerto Rico. Fitopatologia 26: 47.
- Inbar M, Gerling D (2008) Plant-mediated interactions between whiteflies, herbivores and natural enemies. Ann Rev Entomol 53: 431-438.
- Jimenez DR, Yokomi RK, Mayer RT, Shapiro JP (1995) Cytology and physiology of silverleaf whitefly-induced squash silverleaf. Physiol Mol Plant Pathol 46: 227-242.
- Mayer RT, Inbar M, McKenzie CL, Shatters R, Borowicz, (2002) Multitrophic interactions of the silverleaf whitefly, host plants, competing herbivores, and phytopathogens. Arch Insect Biochem Physiol 51: 151-169.
- Inbar M, Doostdar H, Mayer RT (1999) Effect of sessile whitefly nymphs (Homoptera: Aleyrodidae) on leaf- chewing larvae (Lepidoptera: Noctuidae). Environ Entomol 28 : 353-357.
- Mayer RT, McCollum TG, McDonald RE, Polston JE, Doostdar H (1996) Bemisia feeding induces pathogenesis-related proteins in tomato.179-188.
- Blackmer JL, Byrne DN (1999) The effect of Bemisia tabaci on amino acid balance in Cucumis melo. Entomologia Experimentalis et Applicata 93: 315-319.
- Hering EM (1952) Trans Inter Congr. Entomol; 9th Meeting, Amsterdam 1: 507-513.
- Thorsteinson AJ (1960) Host selection in phytophagous insects. Ann Rev Entomol 5: 193-218.
- Hutchinson J (1973) Families of flowering plants. 3rd edition, Oxford at the Clarendon Press: 968.
- De Barro PJ (2005) Bemisia tabaci-from Molecular to Landscape. CSIRO Division of Entomology:1-6.
- Zomlefer WB (1994) Guide to flowering plants. The University of North Carolina Press, Chapel Hill & London: 430.
- Thorne RF (1992) Classification and geography of flowering plants. Bot Rev (Loncaster) 58: 225-348.
- Judd WS, Sanders RW, Donoghue (1994) Angiosperm family pairs: Preliminary cladistics analysis. Harvard Pap Bot 5: 1-51.
- Lotsy (1911) Vortr. Bot. Stammesgesch.3: 658.
- Dahlgren RMT, Clifford HT, Yea PF (1985) The families of the monocotyledonsSpringer Verlag, Berlin: 520.
- Mohan HYR, Wadhi M (1965) Embryology and delimitation of the Acanthaceae. Phytomorphology, 15: 201-205.
- Leng RW(1970) The genera of Acanthaceae in the south eastern United States. J Arnold Arbr 51: 257-309.
- Rettig JH, Wilson HD, Manhart JR (1992) Phylogeny of the Caryophyllales-gene sequence data. Taxon 41: 201-209. Rai BK, Ayachi SS, Rai A (1996) A note on Ethnomedicines from central India. J Econ Taxon Bot Additional Series 12: 186-191.
- Rodman JE (1990) Centrospermae Revisited Part I. Taxon 39: 383-389.