An Overview on Decalepis: A Genus of Woody Medicinal Climbers
Shiwali Sharma and Anwar Shahzad*
Corresponding author: Anwar Shahzad, Plant Biotechnology Laboratory, Department of Botany, Aligarh Muslim University, Aligarh-202 002, UP, India,; E-mail: sharmashiwali@rediffmail.com; ashahzad.bt@amu.ac.in
Plant Biotechnology Laboratory, Department of Botany, Aligarh Muslim University, Aligarh-202 002, UP, India
Citation: Sharma S, Shahzad A. An Overview on Decalepis: A Genus of Woody Medicinal Climbers. J Plant Sci Res. 2014;1(1): 104.
Copyright © 2014 Anwar Shahzad et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Plant Science & Research | ISSN: 2349-2805 | Volume: 1, Issue: 1
Submission: 09/01/2014; Accepted: 10/03/2014; Published: 27/03/2014
Abstract
Decalepis is one of the most important endangered woody medicinal climbing members of “Periploaceace” family. It comprises five species of twining vines and erect shrubs, D. hamiltonii, D. arayalpathra, D. salicifolia, D. khasiana and D. nervosa. Four of the five species of Decalepis are endemic to the Eastern and Western Ghats of peninsular India; the exception, D. khasiana, is geographicallyisolated from the peninsular species, occupying forest areas in the Meghalaya state in the easternmost part of India, Bangladesh, Laos, Myanmar and parts of Southern China. D. hamiltonii is the type species and most widespread of the Indian endemics.
Three species (D. arayalpathra, D. hamiltonii and D. salicifolia) have clusters of numerous fleshy and tuberous roots with a sweet, vanilla-like fragrance due to the presence of 2-hydroxy-4-methoxy benzaldehyde (2H4MB). The tuberous roots of D. arayalpathra and D. salicifolia are moniliform, while those of D. hamiltonii are cylindrical. The roots of D. khasiana are documented as being non-tuberous, andfragrant due to an isomer of vanillin. Four of the five species of Decalepis (all except D. nervosa) are utilized in tribal and traditional Indian and Chinese medicine for the treatment of a wide range of ailments including those of the digestive system, lungs and circulatory system. Presently the three peninsular Indian species of Decalepis are threatened in the wild, and listed as endangered (D. hamiltonii) to critically endangered (D. arayalpathra and D. salicifolia) due to over-exploitation and habitat loss. During last few years considerable efforts havebeen tried to conserve this valuable endangered liana using different strategies of plant tissue culture viz., in vitro shoot regeneration (direct or indirect) and root induction, somatic embryogenesis, hydrogel encapsulation, normal root culture and hairy root culture. The present review provides up-to-date baseline information of all the species of this valuable endangered and endemic medicinal genus forfurther research work.
Keywords: D. hamiltonii; D. arayalpathra; D. salicifolia; D. khasiana; D. nervosa
Abbreviations
BA: N-6-Benzyladenine; 13C NMR: Carbon 13 NMR; IAA: Indole- 3-Acetic Acid; IUCN: The International Union for Conservation of Nature and Natural Resources; GA3: Gibberellic Acid; GC: Gas Chromatography; GC-MS: Gas Chromatography Mass Spectroscopy; 2H4MB: 2-Hydroxy-4-Methoxy Benzaldehyde; 2-iP: 2-IsopentylAdenine; Kn: Kinetin; NAA: α-Naphthalene Acetic Acid; PAA: Phenyl Acetic Acid; PG: Phloroglucinol; 1H NMR: Proton NMR; AgNO3: Silver Nitrate; TDZ: Thidiazuron; TLC: Thin Layer Chromatography; TRIA: Triacontanol; Zea: Zeatin.
Introduction
India is one of the 17 megadiversity countries and has all the 13biomes found in the world, with 3 major hotspots out of a total of 34 [1]. It has been reputed to be a treasure house of a wide range of valuable medicinal and aromatic plants on account of habitant of divers’ climatic conditions. India harbours about 47,000 species of plants of which 17,000 are angiosperms [1]. The International Union for Conservation of Nature and Natural Resources (IUCN) Red List of Threatened species include a total of 560 species from India, out of which 247 species are in the Threatened category. On a global basis, the IUCN has estimated that about 12.5% of the world’s vascular plants, totalling about 34,000 species are under varying degree of threat [2]. However, according to the report of Hamilton [3] an estimated 4,000 to 10,000 species of medicinal plants face potential local, regional, national or global extinction, with subsequent serious consequences for livelihoods, economies and health care systems.
Decalepis is one of the most important endangered woody medicinal climbers. This review is aimed at compiling up-to-date information regarding medicinal and economical properties or related information of all the species of Decalepis over its range of distribution, current status and the role of biotechnology in theconservation of this important genus.
Distribution
Decalepis belongs to the family Periploaceace (an offshoot ofthe Asclepiadaceae). It comprises five species of twining vines/erectshrubs;
• D. hamiltonii (the type species)• D. arayalpathra• D. khasiana• D. nervosa• D. salicifolia
Four of the five species of Decalepis are endemic to the Eastern and Western Ghats of peninsular India; the exception, D. khasiana, is geographically isolated from the peninsular species, occupying forest areas in the Meghalaya state in the easternmost part of India, Bangladesh, Laos, Myanmar, and parts of southern China. All five species occur in hilly to montane regions, in elevations ranging from100-1200 meters [4].
D. hamiltonii is the type species and most widespread of the Indian endemics. It is locally called as “Maredukommulu” or “Nannarikommulu” or “Barresugnadhi” or “Maredugaddalu” or “Madinakommulu” in Telugu, “Makaliber” in Kannada, “Magalikizhangu” in Tamil and “Swallow root” in English. It grows in open rocky slopes and crevices of dry and moist deciduous forests of Karnataka (Hassan, Mysore, Bellary, Tumkur, Kolar), Andhra Pradesh (Kurnool, Chittoor, Nellore, Anantapur, Cuddapah districts) and in Tamil Nadu (Chengalpattu, Coimbatore, Dharampuri, Nilgiri) at an altitude from 300-1200 meters [5]. Though relatively widespread, itspopulations are fragmented and gradually declining due to destructive harvesting of the tuberous roots [6].
The remaining peninsular species, D. arayalpathra, D. salicifoliaand D. nervosa, have more restricted distributions, each occupying a different mountain range in the southern part of the Western Ghats. The south Western Ghats (bordered to the north by the Kalindi river) are extremely diverse floristically, comprising the richest center of endemism in peninsular India, with almost 1200 endemic species [7,8]. Subramanyan and Nayar [9] compared this assemblage of hilltops, bounded to the west by the coastal plain and by the semiarid Deccan plain to the east, to an island chain in respect to the distribution of endemic species. In the Western Ghats, the length of the dry season can range from three to eight months over a distance of only 20 km due to windfall patterns and topography, resulting in the development of small ecological islands with unique floras on the borders of wet evergreen and dry evergreen and deciduous forests [9].
D. nervosa, the northernmost of these narrowly endemic species, is restricted to one of the highest sections of the Western Ghats, the Nilgiris (Blue Mountains in Tamil), which occupy the western corner of Tamil Nadu where it borders Karnataka and Kerala states. D. nervosa has been documented at an elevation of ca. 1500 to 2300 m, on rocky hillsides and exposed areas of deciduous forests nearCoonoor, Kothagiri, and Wellington. The Nilgiris are geographically isolated from the neighboring mountains to the south by the 32-42 km wide Palghat gap, a low pass in the Western Ghats [4].
D. salicifolia and D. arayalpathra occupy similar habitats in hillyand mountainous areas to the south of Palghat gap. D. salicifolia has been documented in three localities, the Anamalai Hills (Kerala and Tamil Nadu), Nelliampathy (Kerala) and Marayoor (Kerala) forests. The Anamalai Hills (Elephant hills in Tamil Nadu) straddle Kerala and Tamil Nadu states just south of Palghat gap and are known for their abundant wildlife and under consideration by the UNESCO World Heritage Committee for selection as a World Heritage Site. Further south, D. arayalpathra occupies rocky hill slopes on the Agasthiyar hills (Kerala) and within Kalakkadu Mandanthurai tiger reserve in Tamil Nadu [4].
D. khasiana was first described from a specimen collected at an altitude of 900-1200 m, in the Khasia hills in Eastern India. It has since been documented in Laos, on the Tran-Ninh plateau in the area of Xieng-Khowang, at an altitude of 800-900 m [10], in the Tamabil- Jaflong hills of the Sylhet district in Bangladesh, and in the Yunnan, Guizhou, and Sicuan provinces of China, in bush land forests and in Myanmar [4].
Taxonomic History
The taxonomic history of Decalepis began with the naming of D. hamiltonii by Robert Wight and George Arnott Walker-Arnott in Wight’s “Contributions to the Botany of India” (1834), based on a specimen collected by Wight in Madras, India [11]. Walker-Arnott, who was collaborating with Wight, upon observing what he referred to as “a double row of scales” (referred to in this study as “corona lobes” and “interstaminal nectaries”), placed the new species in its own genus, Decalepis (Greek deca=ten, lepis =scale) [11]. Wight mistakenly referred to the specimen in an earlier, unpublished version of his manuscript as being from the herbarium of the Scottish physician and botanist Francis Hamilton; Walker-Arnott chose to honor Hamilton with the specific epithet hamiltonii despite the fact that the specimen was from the Madras Herbarium and not from Hamilton’s collection [4].
Decalepis was monotypic until Venter and Verhoeven [12] synonymized the monotypic Indian genera Baeolepis Decne. ex Moq. and Janakia J. Joseph & V. Chandras. with Decalepis, followed in 2001 by Utleria Bedd. ex. Benth. & Hook. f., also monotypic and endemic to India, on the basis of “ovate-like corona lobes, an obconical style-head with flat apex containing translator hollows, and pollen in pollinia” [12,13]. Results of phylogenetic analysis (excepting D. nervosa, for which molecular data were unavailable) tentatively support this grouping, with the addition of D. khasiana, which Venter and Verhoeven [13] had moved previously from the monotypic Stelmacrypton Baill. to Finlaysonia along with several other monotypic and bitypic Asian genera, on the basis of similarities in floral form i.e., the gynostegium exposed from a rotate or saucershaped corolla, staminal feet swollen and these topped by corona lobes and pollen tetrads fused into pollinia (most Periplocoideae have pollen in the form of free tetrads). Subsequent molecular studies have demonstrated Finlaysonia sensu [13] to be polyphyletic, and the characters used to support that grouping to be pleisiomorphic [14].
Habit and Habitat
Three species of Decalepis (D. hamiltonii, D. nervosa and D. khasiana) are scandent to climbing twiners, the commonest growth habit among Periplocoideae [13], while two (D. salicifolia and D. arayalpathra) are erect shrubs, a condition that has evolved numerous times within the subfamily and is generally correlated with arid environments or those with seasonal water shortages [15]. Both D. salicifolia and D. arayalpathra are narrow endemics of poor soils in rocky hill slopes and rock fissures in deciduous and evergreen forests, at elevations above 600 m. However, the vining species of Decalepis tend to occupy habitats similar to those of the erect species, particularly the narrow endemic D. nervosa.
Botanical Description
Inota (2006) provided a comparative taxonomic detail of all the five species of Decalepis on the basis of 66 herbarium specimens from BM, CAL, E, K, LM, MO, NY, P and US [16] Figure 1.

Figure 1: Different plant parts of D. hamiltoniiA. Cylindrical tuberous roots excised from a five year old plant.B.Dry follicle with seeds.C.Seeds with silky hairs.D.Fresh seeds.E.Complete plant of D. hamiltonii planted in loamy soil-6 month old.
i. Roots: Three species have clusters of numerous, fleshy, tuberous roots with a sweet, vanilla-like fragrance: D. arayalpathra, D. hamiltonii and D. salicifolia. The tuberous roots of D. arayalpathra and D. salicifolia are moniliform, while those of D. hamiltonii are cylindrical. The roots of D. khasiana are documented as being non-tuberous, and fragrant due to an isomer of vanillin that is also present in the other fragrant-rooted species. No references were found pertaining to the root morphology or chemistry of D. nervosa [4].
ii. Stems: The stems are woody and reddish brown when young, tend to become purplish when older [12]. Lenticels are common on older branches. The stems of D. salicifolia are somewhat succulent, becoming quite wrinkled when dried and the older stems are covered with a grayish wax [4].
iii. Leaves: The leaves are opposite, decussate, petiolate and exstipulate. In D. salicifolia the leaves are restricted to the ends of the young branches and clustered at the shoot apices, where they sometimes appear to be alternate or whorled due to reduced internodes at growing shoot tips. The lamina rangesfrom 2.9-11.5 cm long in all but D. salicifolia, a species having leaves that may be up to 19.9 cm long, and narrowly lanceolate, in contrast to the elliptic to ovate or obovate leaves of the other species. Leaf apices are acuminate, obtuse or rounded, and in D. hamiltonii sometimes retuse. The acute to obtuse leaf basesin D. nervosa distinguish it from the other species, which have decurrent leaf bases. The lamina is generally flat, but in D. hamiltonii it is curved downwards abaxially, particularly at the tip, where it is recurved-falcate [4].
iv. Inflorescence: The inflorescences are sparsely to multiple branched, few- (up to 10 in D. khasiana and D. arayalpathra; 15 in D. hamiltonii) to many- (up to 50 in D. salicifolia; 80 in D. nervosa) flowered axillary cymes. Branching is monochasial throughout in D. hamiltonii, D. arayalpathra and D. salicifolia;in D. nervosa the inflorescences branch dichasially proximally and monochasially distally, and branching is dichasial throughout in D. khasisana [4].
v. Flower structure and terminology: The flower buds of Decalepis are ovoid; the corollas slightly twisted in bud and with dextrorsely imbricate aestivation. The flowers are perfect, actinomorphic, and pentamerous. D. arayalpathra has the smallest flowers compared to other species, almost 2.5 mm long and 4.0 mm wide at anthesis. The other species have flowers to 3.2 mm long and 5.3 mm wide, with the exception of D. khasiana, which has flowers that are over twice the size of most i.e., 6.5 mm long and 10.0 mm wide [4].
vi. Calyx: The sepals are free, ovate to obovate (oblong-ovate in D. hamiltonii), and 0.7-1.8 mm long by 0.6-1.5 mm wide, with an acute to obtuse (to rounded in D. salicifolia) apex and an irregular to entire, the margin occasionally sparselyciliate apically (but margin hairs lacking in D. khasiana). The sepals of D. salicifolia flowers are more broadly obovate than those of other species, and thinner, lacking an obvious midrib. Brownish oblong to triangular colleters are located singly or in pairs at the inner bases of overlapping margins of adjacentsepals, but these are occasionally absent in D. hamiltonii [4].
vii. Corolla: The corolla is shallowly campanulate (to rotate inD. khasiana), with ovate to oblong corolla in all species but D. khasiana, with acute to obtuse apices. In most species the corolla is cream to yellow or greenish, but in D. nervosa and D. khasiana the flowers are rose to dark purple. The corolla lobes of D. nervosa flowers are thicker than in most species, with a tuberculate adaxial surface (smooth in other species); deep grooves radiate from the inner corolla tube adjacent to the base of each corolla lobe, oriented below the pollen translators and extending ca. half way to the lobe apex. Interestingly, this species lacks the tubular nectar lobes present in others of the genus, the groove acting in tandem with the shelf-like nectaries to form nectar collecting pockets [4].
viii. Corona: The corona lobes alternate with the corolla lobes, emerging just above the filaments and in most cases fused laterally to the upper corolla tube to the level of the corolla sinuses, and free above that. In D. hamiltonii, the corona lobes are free from the corolla tube (i.e., attached only at base). In D. nervosa, D. salicifolia and D. arayalpathra, the lobes are, respectively, rectangular, oblong or triangular ovate, with rounded to obtuse apices, and ca. 0.2-0.57 mm long androughly as wide. D. khasiana has triangular lobes about 1.4 mm long and 0.8 mm wide at the base, with an acute apex, and the corona lobes of D. hamiltonii are lanceolate and ca. 1.1 mm long by 0.6 mm wide. The corona lobes of D. nervosaare uniquely minutely tuberculate abaxially, with an irregular to lobed apex [4].
ix. Androecium: The stamens are inserted on the corolla just below the corona lobes, from 0.10 to 0.35 mm (1.0 mm in D. khasiana) below the corolla sinuses. The anthers are ovate to oblong-ovate (deltoid in D. nervosa), from 0.5 to 0.6 mmlong, excluding the connective appendage (1.0 mm long in D. khasiana). The filaments (absent in D. nervosa) are generally short (ca. 0.2 mm long) but longer (1.0 mm) in D. khasiana, and dilated at the base in D. arayalpathra, D. salicifolia, andD. hamiltonii (in the last species they are also adnate to the corona). Swollen staminal feet are present on the lower corolla tube just below the filaments in those species lacking dilated filament bases. In most species the stamens are glabrous, but D. khasiana has hairs scattered along the abaxial anther connective and connective appendage [4].
x. Pollen and Translators: Pollen is in the form of tetrads that are agglutinated into pollinia at anthesis and deposited onto spoon-shaped translators. The tetrads are elongate and uniplanar, generally elliptic to obovate, and in most speciesthe individual grains of the tetrads are arranged into a mix of rhomboidal (the most common shape), t-shaped, and linear forms. In D. nervosa only rhomboidal tetrads were observed. D. hamiltonii is unique in having predominantly linear tetrads (occasionally rhomboidal). The distal walls of the tetrads forming pollinia are in aperturate. Pollen translators are spoon-like and distally dilated into a broad obcordate concave receptacle with an emarginate apex. The receptacle has a longitudinal groove on the abaxial surface, effectively forming two shallow cups in which pollinia are deposited. The viscidium is discoid. D. khasiana and D. hamiltonii have the largest translators, with receptacles 0.6-0.4 mm long and stipes ca. 0.2 mm long. In other species the translator receptacles are 0.15-0.25 mm long, the stipes 0.05 to 0.1 mm long [4].
xi. Nectaries: In Decalepis the nectaries are located between adjacent pairs of stamens (interstaminal) and just below them, and in most species they are tabular in form, with the exception of D. nervosa, in which they are in the form of a low ledge. The tabular nectar discs are flattened in cross section and fleshy, and oriented upwards, so that together with the adjacent dilated filament bases (or staminal feet in D. khasiana), they surround the style branches, causing thenectar to collect in the upper corolla tube below the pollen translators, thus attracting pollinators towards a position that is favorable for pollen transfer. The ledge-like nectar discs of D. nervosa similarly surround the style (alternating with swollen staminal feet), the nectar collecting in the deep grooves in thecorolla that are oriented just below the pollen translators. The tabular nectaries are fleshy and similar in shape to the corona lobes (but shorter, from 0.2-0.6 mm long) in D. hamiltonii, D. salicifolia, and D. arayalpathra. In D. khasiana the lobes are rectangular with rounded edges, in contrast to its triangular corona lobes [4].
xii. Gynoecium: The ovaries are bicarpellate, subglobose, and half-inferior, and vary from 0.8 to 1.2 mm long. The compound, more or less terete styles are from 0.2-0.3 mm long (to 0.7 in D. hamiltonii), becoming united and dilated atthe apex to form a pentagonal stylar head, which is obscurely 2-beaked at the center with depressions on the upper surface where the pollen translators (a hardened secretion of the stylar head) are formed. In Periplocoideae the upper surface of thestylar head is usually domed and of the species of Decalepis, the stylar head of D. khasiana, which is deltoid in longitudinal section, is most typical for the subfamily. In other species of Decalepis, the stylar head is much shallower, in all cases wider than tall. In D. hamiltonii and D. nervosa the upper surface is concave, and in D. salicifolia and D. arayalpathra it is broadly convex [4].
xiii. Fruits: The fruits of Decalepis, like other Periplocoideae, arepaired, divaricate follicles. In Decalepis they are more or less cylindrical (D. hamiltonii with a median groove along the suture), narrowing distally towards an acute apex, and 3.5-7.5 mm long. The endocarp is waxy, bright yellow and ca. 0.2 mmthick. The mesocarp is relatively thin in most species (ca. 0.4 mm), but markedly thicker in D. hamiltonii (1.5 mm). The exocarp is light to dark brown, and in D. hamiltonii and D. salicifolia extremely waxy, and wrinkled when dry. The fruitsof D. khasiana are not known [4].
xiv. Seeds: The seeds are elliptic to obovate and flattened (curved along the longitudinal axis towards the funicular side in D. nervosa), with a coma of white to yellowish hairs up to 2.9 cm long at the micropylar end. Coma hair color varies significantly within species and is not taxonomically informative. The seedsare slightly keeled at the point of attachment to the placenta, and in D. hamiltonii and D. nervosa the surface is smooth or somewhat wrinkled on the funicular side. Testa color ranges from dark red-maroon to dull black, and varies within the species. Seeds of D. khasiana are not known [4].
Cultivation
For D. hamiltonii, loamy soils are well suited for the growth but soils intermixed with stones encourage the production of long thick roots. The length and thickness of the roots mostly depends on the soils profile rather than the environment [17]. However the information regarding cultivation for other species of Decalepis is not well documented anywhere.
Propagation
All the species of Decalepis can be propagated from seeds as well asby means of stem cuttings. For commercial purpose the first method is preferred. As the direct sowing of seeds in the field does not give good results, seedlings are raised in the nursery before transplanting in the field. Collection of seeds from February-March is preferred to achieve higher viability percentage. Because of short viability, the fresh seeds are to be used. The seeds can be raised either in the nursery beds or in polythene bags. Germination starts after about 2 weeks andmay continue for another week. Three months old seedlings having 4-6 leaves are transplanted at a distance of 60 cm × 60 cm in field. Generally, That sentence will be replaced by this: the crop is cultivated under rain-fed conditions [17]. Seeds of D. hamiltonii were found to germinate soon after early monsoon showers in May on soil floor and rocky areas which are rich in litter and other organic matter. In nature, seed germination rate is 13% out of which seedling establishment rate is 48%. Thus, the overall seedling establishment rate in relation to total seeds sown is only 6% [18]. Anandalakshmi and Prakash [19] have studied the viability and germination of D. hamiltonii seeds under in vitro conditions. They have suggested that the use of moist filter papers was better substrate for germination (83 to 98%) than sand (15 to 28%) after pre-soaking of seeds in hot water (60°C) for 24 hours. Pretreatment study coupled with microscopic studies indicated that about 14% of seeds were found to be hard seeded. Seeds could tolerate desiccation up to 10% moisture content (65% germination) while nil germination was recorded at 5% seed moisture content. The seeds were found to be chilling sensitive, thus showing total mortality at 10°C. Rapid depletion of seed metabolites is indicative of short viability. Based on the storage behaviour, D. hamiltonii seeds could be classified as intermediate seeds. From seed perspective, hard seed coat, need for a well aerated substrate and short viability could be the likely impeding factors for inadequate regeneration of the species in wild. Saini and Giridhar [20] found that exposing seeds to 0.3% H2O2 resulted in effective (94%) seed germination in D. hamiltonii. While Vedavathy (2004) reported 100% germination of immature zygoticembryos when placed on MS medium supplemented with GA3 (0.05 ppm), BA (1.0 ppm) and TDZ (1.0 ppm).
Harvesting
The roots of D. hamiltonii come to maturity in about 12-14 months after planting depending upon the soil and climatic conditions. The roots can easily be harvested by digging around the plant leaving the central core of the root and stem for regeneration. Some plants could be used to harvest roots up to 4-5 years, as vigorous regeneration and growth resumed even with a single left over root. Each root is 5-10 cm in diameter and 4-10 roots regenerate from the rootstock. A 2-3 year old plant produces 15-20 kg of roots, while one year old plant produces 1-2 kg of roots. Roots are harvested during summer months mostly by the Yanadi tribe of Chittoor district and it is the main source of income to them until the agricultural works resumes.
Chemical constituents
The roots of D. hamiltonii have been reported to contain aldehyde, inositols, saponins, amyrins, lupeols [21,22]. Morphologically as well as chemically the plant resembles African liana called Mondia whitei. Both have similar ethno botanical uses and the presence 2-hydroxy-4-methoxy benzaldehyde (2H4MB), an isomer of vanillin is reported from the volatile oils (96%) [23]. Further GC-MS analysis of this oil showed, along with the major component, the presence of benzaldehyde (0.017%), salicylaldehyde (0.018%), methyl salicylate (0.044%), benzyl alcohol (0.016%), 2-phenylethyl alcohol (0.081%), ethyl salicylate (0.038%), p-anisaldehyde (0.01%), and vanillin (0.45%) in minor quantities which are biologically significant [23]. Hydro-distillation of D. hamiltonii roots yielded an essential oil (0.33% v/w) that contained 2H4MB (37.45%), 2-hydoxybenzaldehyde (31.01%), 4-O-methylresorcylaldehyde (9.12%) and benzyl alcohol (3.16%) as major constituents, with aromatic aldehydes constituting the main fraction of its roots’ essential oil [24]. Thangadurai et al. [25] isolated two other compounds from the root extract that potentially account for its insecticidal and anti-microbial properties: α-atlantone (2.06% v/w of oil), commonly used to control household insects and nontoxic to mammals, and β-pinene (2.01%), which is used as an insecticide and disinfectant. Whereas, Zhang et al. [26] isolated four new pregnane glycosides (Stelmatocryptonoside A, B, C and D) from the stems of D. khasiana; and four species of sariva (D. hamiltonii, Cryptolepis buchananii, Ichnocarpus frutescens and H. indicus) were shown to have potent anti-inflammatory properties [27].
A gas chromatographic (GC) procedure was developed by Nagarajan and Rao [28] for the assay of 2H4MB in both fresh and dried roots of D. hamiltonii from different origins. They used benzyl butyrate as an internal standard. Among the methods tried, steam hydrodistillation was found most suitable for extraction of the volatile oils. The quantity of this aromatic compound varied from 0.03 to 0.54% (Nagarajan and Rao 2003). In addition 4-hydroxyisopthalic acid, 14-aminiotetradecanoic acid, 4-(1-hydroxy-1-methylethyl)-1-methyl- 1,2-cylohexane diol, 2-(hydroxymethyl)-3-ethoxybenzaldehyde, 2,4,8-trihydroxybicycle (3,2,1) octan-3-one, bis-2,3,4,6-galloyl-α/β- D-glucopyranoside, bornerol and ellagic acid have been identified in D. hamiltonii root [29,30].
Traditional uses
Four of the five species of Decalepis (all except D. nervosa) are utilized in tribal and traditional Indian and Chinese medicine for the treatment of a wide range of ailments including those of the digestive system, lungs and circulatory system [31-33,26]. The principal organs used are the tuberous roots, which are fragrant and sweet tasting (fragrant but non-tuberous in D. khasiana), with a vanilla-like taste and odor [34,33,28,35]. D. hamiltonii, the type species, is the most widespread and utilized species in the genus.
The roots of D. hamiltonii are also used in folk medicine and in Ayurvedic preparations [36]. Dried roots are available in Indian drug stores and local tribal markets. Ancient tribes in the Western Ghats of India use its roots particularly for inflammation [37]. These people procure and habitually carry the roots with them and chew the same whenever the digestion may seek relief. Besides treating indigestion, the roots have also been used locally to stimulate the appetite and to relieve flatulence and to act as a general tonic [17]. Tuberous roots are used as a flavoring principle [21] and a blood purifier [31,38]. Roots are used to cure dysentery, cough, bronchitis, leucorrhoea, uterine hemorrhage, skin disease, fever, indigestion, vomiting, chronic rheumatism, anemia and blood diseases. They are also consumed as pickles and as a popular cool drink in the forest areas of the Eastern and Western Ghats, known as Nannari which has a cooling effect without any toxic effects on human beings [39,17,40]. It finds its use as culinary spice because of its high priced aromatic roots and marketed on a large scale [18]. Root extract can be used not only as food preservative (to replace for the toxic butylatedhydoxy anisole and butylatedhydoxy toluene currently under use) but also can be used in the preparation of neutraceuticals and pharmaceutical products [41]. The roots of D. hamiltonii are little bitter and then sweet. It is so characteristic with a familiar lingering after taste and smell of vanillin, the substance that is in Vanilla planifolia, an orchid used in ice-creams, chocolates, drinks etc. Although vanillin has been synthesized since 1874 natural source of this flavoring are still in demand and the roots of Decalepis species can be used as substitute for vanillin.
D. arayalpathra was discovered relatively recently, and described as Janakia arayalpathra [42]. Pushpangadan et al. [31] discovered during an ethno-medico-botanical investigation that though relatively new to science, D. araylpathra has long been recognized by the Kani tribe of Kerala, they consider it an important medicinal plant, as reflected in the vernacular name ‘amrithapala’, which translates as ‘the plant which gives the milky ambrosia or nectar of immortality’(amritha=ambrosia or ‘nectar of immortality’, pala=milk, likely referring to the abundant white latex present in all plant parts). They make use of juice obtained from the tuber as a remedy for peptic ulcer, as a rejuvenating tonic, and to cure for external cancers [32]. In all other species, D. salicifolia has been known to science since its description in the 19th century; its ethnobotanical use by the seminomadic Kadars, Malasars, and Muthuvans tribes was discovered only recently. The tubers of ‘Mahali kizhangu’ (Mahalekshmy=goddess of wealth, kizhangu=root tubers) are hung from the front of huts to bring good fortune, and pickles made from the roots are used to treat colic, bleeding ulcer, tuberculosis, asthma, and skin diseases [33]. Similarly, all parts of D. khasiana including its fragrant, vanilla-scented roots are used in Chinese folk medicine, for the treatment of bronchitis, cough, stomach and rheumatic aches, and influenza. No records ofethnobotanical use were found for D. nervosa.
Medicinal Importance
Numerous recent investigations have tested the medicinal properties of the different species of Decalepis supported the veracity of various traditional ethnomedicinal claims. As compared to other species, D. hamiltonii has been much evaluated for its medicinal properties as below;
i. Anti fungal activity: The antifungal activity of aqueous extract of D. hamiltonii was evaluated at different concentrations by poisoned food technique against 8 species of Fusarium, 10 species of Aspergillus, 3 species of Penicillium, 2 species of Drechslera and Alternaria alternata. [43]. Mohana et al. [44] found that the antifungal active principle is a phenolic compound, 2H4MB. In vitro antifungal activity assay of different concentrations of 2H4MB isolated from D. hamiltonii against 6 important seed-borne fungal pathogens, namely, A. alternata, Drechslera tetramera, Fusarium oxysporum, Fusarium proliferatum, Pyricularia oryzae and Trichoconis padwickii isolated from paddy seeds revealed that, the compound 2H4MB showed significant antifungal activity. Among the fungi tested, F. proliferatum showed the highest inhibitory activity, whereas P. oryzae showed least inhibitory activity [44].
ii. Antibacterial activity: Thangavel et al. [45] studied antibacterial potential of crude petroleum ether extract of leaf callus tissue of D. hamiltonii against 5 bacterial pathogens using agar well diffusion method. Among the tested bacterial strains, inhibitory activity of the callus extract was minimum against Klebsiella pneumoniae (5 mm) and followed by Proteus vulgaris (6 mm). Maximum inhibitory activity was observed against Salmonella typhi (11 mm). From this observation it was evident that phytochemical principles which are responsible for the curative (antibacterial) activity of D. hamiltonii could have a constant expression pattern in a specialized set of cells even after their rapid division. Later on Devi and Latha [46] also reported antibacterial activity of various root extracts of D. hamiltonii against E. coli, K. pneumoniae, S. typhi, Proteus mirabilis, Vibrio cholerae, Shigella sonnie, Serritias species, S. aureus and Bacillus subtilis by disc diffusion method.
iii. Insecticidal activity Several studies have been conducted to evaluate the insecticidal activity of D. hamiltonii against 3 coleopteran stored product pests,viz., rice weevil, the lesser grain borer and red-rust flour beetle [24,47].Further, the residual deposits of 2H4MB has been assayed for contact toxicity on rice weevil (Sitophilus oryzae), Rhyzopartha dominica and Tribolium castaneum. Thus, the aromatic chemicals of D. hamiltonii have been established with insecticidal and pesticidal properties.
iv. Anti-oxidant activity: Free radicals are implicated in the etiology of several diseases, such as cancer, antherosclerosis, diabetes, neurodegenerative disorder and aging. Antioxidants prevent the damage to macromolecules and cells by interfering with the free radicals. Natural products containing antioxidants from plants are believed to modulate oxidative stress and to prevent or delay degenerative disorders [48]. Harish et al. [29] characterized 6 pure compounds with strong antioxidant properties from the methanolic extract of D. hamiltonii roots by using MS, 1H NMR, 13C NMR and two-dimensional NMR spectroscopic techniques and including a novel antioxidant molecule, bis-2,3,4,6-galloyl-α/β D glucopyranoside (aptly named decalepin) from D. hamiltonii root extracts. Srivastava et al. [29] also reported antioxidant potential of the methanolic and aqueous extracts of D. hamiltonii roots. Murthy et al. [49] found that antioxidant activity was associated with 2H4MB. They subjected different parts of aromatic roots viz., whole tuber, peel, tuber without peel and medullary portion. Maximum antioxidant activity was found with peel extract followed by medullary Portion [49]. Later on, Srivastava et al. [50] isolated another antioxidant compound from the aqueous extract of the roots of D. hamiltonii and identified it as ellagic acid (EA), based on NMR and MS studies. Anti-typhoid activity Kumuda et al. [51] studied in vitro anti-typhoid activity of various roots extracts of D. hamiltonii. The powdered roots were extracted with various solvents viz., petroleum ether, chloroform, and ethyl acetate by Soxhlet extraction method and the extracts were tested on typhoid causing organisms. Among these, extracts of petroleum ether and chloroform showed a significant activity against S. Typhi (ATCC 14028), S. paratyphi A and S. paratyphi B respectively on comparison with Ciprofloxacin [51].
v. Ulcer preventive activity Srikanta [30] reported a pectic polysaccharide from D. hamiltonii roots containing a sulphonamide group and phenolics as an effective antiulcer compound in vivo. They envisaged a multi-potent role for this phenolic polysaccharide in the upregulation of mucin, antioxidant levels, modulation of oxidative status, inhibition of H+, K+-ATPase activity against swim and ethanol stress induced ulcers in experimental animal models, in addition to its ability to inhibit Helicobacter pylori. Roots containing phenolics at 0.12 g GAE/g, prevented stress-induced gastric ulcers in animal models by 80-85%. Histopathological analysis revealed protection to the disrupted gastric mucosal layer and epithelial glands. SRPP also inhibited H+, K+-ATPase in vitro, at an IC50 of 77 μg/mL as opposed to that of 19.3 μg/mL of Lansoprazole and H. pylori growth at Minimum Inhibitory Concentration (MIC) of 150 μg/mL. In addition, free radical scavenging (IC50-40 μg/mL) and reducing power (3200 U/g) activities were also observed. They also concluded that SRPP is non-toxic as opposed to other known antiulcer drugs, and therefore may be employed as a potential alternative for ulcer management.
vi. Anti-diabetic activity: Sumalatha et al. [52] evaluated the anti-diabetic activity of extracts of the D. hamiltonii in alloxan induced diabetic rats. The fall of blood glucose levels after administration of aqueous, methanol and petroleum ether extracts at a dose of 200 mg/ kg body weight were found to be 69.43, 62.04, and 49.61%, respectively, after 4 hours of oral administration. At the same dose the acute oral administration of aqueous extract showed significant decrease of blood glucose loaded normoglycemic rats. In vitro glucose uptake studies suggest that D. hamiltonii root extracts has direct insulin like effect which can enhance the peripheral utilization of glucose by rat hemi diaphragm.
vii. Antitumor activity: Cancer is a major aliment that affects several organs and chemothaerapy is widely practiced to treat the disease. Chemotherapy with or without radiation, although effective for many cancers, is accompanied by severe side effects due to the toxicity of the anticancer drugs. Several anticancer drugs are derived from plants such as taxol, cycolphosphamide, vincristine, vinblastineand there is great interest in the search for newer and safer drugs from plants. In this regard, Zarei and Shivanandappa [53] have shown the antitumor effect of D. hamiltonii root extract (DHA) against Ehrlich Ascites Tumor (EAT) cells in mice and compared its effect with that of the anticancer drug, cyclophosphamide (CP). Treatment of EAT cell bearing mice with aqueous extract of D. hamiltonii (50 mg/kg and 100 mg/kg b.w.) showed a significant reduction in ascites tumor volume, tumor cell counts and increased median survival time (MST). EAT cells induced alterations in hematological profile and the serum marker enzymes aspartate abd alanine transaminases (AST, ALT), alkine phosphatase (ALP) and lactate dehyrogenase (LDH) were restored by DHA treatment. The antitumor effect of DHA was comparable to that of the anticancer drug cyclophosphamide suggesting the anticancer potential of the root extract of D. hamiltoniii [53].
Causes of its Extinction and Need of Micropropagation
In traditional Indian medicine, an estimated 90% of the species used for plant-based medicines are collected from wild populations, and roots are the plant organ most commonly used, resulting in destructive harvest since the whole plant must be pulled from the ground [35]. Presently the three peninsular Indian species of Decalepis are threatened in the wild, and listed by IUCN (International Union for Conservation of Nature) as endangered (D. hamiltonii) to critically endangered (D. arayalpathra and D. salicifolia) due to overexploitation and habitat loss. Destructive harvesting of the narrow endemics D. arayalpathra and D. salicifolia, particularly vulnerable given their limited distributions, is escalating as their medicinal properties become known [54] and even the relatively widespread D. hamiltonii faces threats due to its increasing use as a substitute for the important Ayurvedic medicinal plant Hemidesmus indicus [55]. In D. hamiltonii, extended flowering pattern, self-incompatibility, pollinator limitation, absence of seed dormancy, abortion of a considerable percentage of seedlings prior to establishment are contributing factors for the regulation of its population size [56]. Moreover, the absence of any organized cultivation of this plant [57] calls for immediate conservation measures. Recent efforts aimed at meeting the growing demand for these plants and their useful products while at the same time ensuring the protection of wild populations include clonal propagation techniques using nodal/axillary bud explants, which have been developed and optimized for each of the three ethnomedically important peninsular species [55,58-61,54]. The Foundation for Revitalization of Local Health Traditions (FRLHT), a non-governmental organization based in southern India, has been monitoring populations of D. hamiltonii and established Medicinal Plant Conservation Areas focused on the in situ conservation of D. arayalpathra and D. salicifolia.
Role of Biotechnological Tools for ex vitro Conservation
In vitro propagation of rare and threatened plants is generally undertaken to multiply and conserve the germplasm especially when population numbers are low in wild, difficult to regenerate by conventional methods and where population has decreased due to over exploitation by destructive harvesting. The modern technology of micropropagation provides numerous advantages over conventional propagation methods like mass production of true-to-type and disease free plants of elite species in a highly speedy manner irrespective of the season within smaller space and tissue source. During last few years considerable efforts have been tried to conserve this valuable endangered liana Table 1. Following strategies of plant tissue culture have been used for efficient propagation and conservation;
i. In vitro shoot regeneration (direct or indirect) and root inductionii. Somatic embryogenesisiii. Hydrogel encapsulationiv. Normal root culturev. Hairy root culture
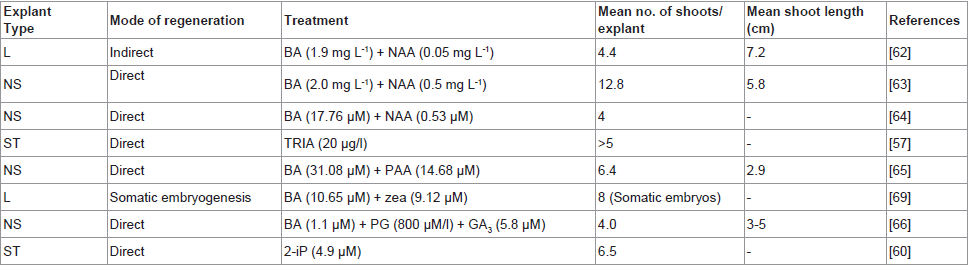
Table 1: In vitro shoot regeneraion using different explants of D. hamiltonii (chronological order).
In vitro shoot regeneration (direct or indirect) and rootinduction
D. hamiltonii: For the first time, George et al. [62] studied in vitro growth and development of multiple shoots from leaf calli of D. hamiltonii. They used “Response Surface Methodology” (RSM) to evaluate the effect of BA, NAA and sucrose on multiple shoot induction and to develop an efficient micropropagation system. The optimum combination for maximum shoot regeneration (4.4 shoots) was 1.9 mg L-1 BA and 0.05 mg L-1 NAA. Shoot number exhibited a positive linear relationship with BA concentration and was maximum at the highest levels of BA, while sucrose had no significant effect. However, maximum shoot length was observed when sucrose concentration was maximum (3%) in the optimal medium. Bais et al. [63] used axillary bud for micropropagation and achieved maximum shoot number (12.8 shoots per culture), shoot length (5.8 cm) and number of leaves (16.4) on MS medium supplemented with BA (2.0 mg L-1) and NAA (0.5 mg L-1) after 30 days of incubation. Similarly, Anitha and Pullaiah [64] used nodal segments and reported maximum of 4 shoots per explant on MS medium supplemented with 17.76 μM BA and 0.53 μM NAA. Reddy et al. [57] studied the effect of triacontanol (TRIA) on shoot multiplication of in vitro derived shoot tips. Among different concentrations (2-20 μg L-1) of TRIA used, 20 μg L-1 TRIA was found to be most effective for shoot proliferation. While Giridhar et al. [65] found that the addition of phenylacetice acid (PAA) had a significant effect on shoot multiplication through nodal segments. The maximum number of shoots per culture (6.4 shoots) was produced on MS medium supplemented with 31.08 μM BA and 14.68 μM PAA, while the longest shoot length (4.5 cm) and nodes were obtained on MS medium supplemented with 22.2 μM BA and 14.68 μM PAA. Shoot subcultured on MS medium supplemented with 22.2 μM BA and 14.68 μM PAA elongated along with secondary shoot formation.
Similarly Gururaj et al. [66] found that phloroglucinol (PG) had synergistic effect on shoot multiplication though nodal segments when added with BA and GA3. Maximum number of shoots per culture was observed on MS medium containing 1.1 μM BA, 5.8 μM GA3 and 800 μM PG. Subculturing of the shoots onto MS medium containing optimum concentration of BA (5.6 μM), PG (200 μM) and TRIA (0.11 μM) produced elongated shoots along with secondary shoot formation. Later on, Giridhar et al. [67] compared the influence of different cytokinins (2-iP, BA, Kn, TDZ and zea) on shoot multiplication through shoot tips. Maximum number of multiple shoots (6.5) was noticed on MS medium supplemented with 4.9 μM 2-iP. Further elongation of shoots and adventitious shoot formation was obtained on MS medium with 2.5 μM 2-iP and 0.3 μM GA3 (Table 1).
Giridhar et al. [60] reported the improvement of 2H4MB of micropropgated plants under TRIA treatment. The microppropgated plants were raised from nodal explants on MS medium supplemented with 2.0 mg/l and 0.5 mg/l NAA in stage I and MS medium supplemented with 2.0 mg/l IBA in stage II by following the method reported earlier by Bais et al. [63]. They were then hardened in a green house for 2 months. These micropraogated plants of 12-15 cm shoot length were used in experiments. They observed improved plant growth and yield of tubers with enhanced flavor compound, by using TRIA soil drenching. The administration of TRIA (0.228 μM) soil drenching twice by giving second treatment with a gap of 60 days after initial treatment, showed fast sign of growth and good yield of tubes with enhanced flavor content. The GC-MS profile of 2H4MB in tubers showed significant improvement over controls (1.5 times more), which could be useful for various commercial application.
Giridhar et al. [68] analysed the production of 2H4MB in roots of tissue culture raised and acclimatized plants of D. hamiltonii. They used axillary buds excised from field grown plants to initiate multiple shoots on MS medium supplemented with 2 mg/l BA and 0.5 mg/l IAA. Profuse rooting was achieved when actively growing shoots were cultured on MS medium supplemented with 1.0 mg/l IBA. Regenerated plants were grown successfully in the plains, in contrast to wild growth in high altitudes and rocky crevices of hilly regions. Roots of different sizes from one-year-old tissue culture raised field grown plants had the same profile of 2H4MB as that of wild plants. A maximum of 0.14% and 0.12% 2H4MB was produced in roots of one year old tissue culture derived plants and greenhouse grown plants respectively.
The success of in vitro regeneration relies on an efficient rooting in regenerated shoots and their subsequent acclimatization which is often problematic in some plant species. Bais et al. [63] investigated in vitro rooting of D. hamiltonii by using silver nitrate (AgNO3). They found that medium comprising of MS salts with IAA (2.88 μM), induced poor rooting and root emergence did not occur until after 25 days. The resultant roots were stunted. However, the addition of 40 μM AgNO3 improved root initiation and elongation. The promotive effects of AgNO3 on rooting may result from inhibition of ethylene action. It was noticed that the addition of ethephon to the rooting medium, excessive callusing was observed in explants in all the treatments. Addition of 40 μM AgNO3 to ethephon containing medium resulted in improvement in root initiation and elongation. Ethylene production was monitored in all the treatments with IAA/ AgNO3/ethephon and it was observed that the treatment with IAA (2.88 μM) alone showed a greater increase in ethylene production when compared with AgNO3, ethephon and their combinations.
For efficient rooting of in vitro-derived microshoots by using phloroglucinol (PG), activated charcoal and CoCl2 was investigated by Reddy et al. [58]. Transfer of in vitro-derived shoots to MS medium supplemented with 8.8 μM IBA and 1.43 μM IAA resulted in root induction. IBA was found to be a good root-promoting agent, dipping of explants in 4.4 μM IBA for 30 min and subsequent inoculation on MS basal medium was also beneficial for root induction Reddy et al. [58]. While, Anitha and Pullaiah [64] reported a maximum of 2 to 3 roots on MS medium containing 16.11 μM NAA. Whereas Reddy et al. [57] found that TRIA at 5 and 10 μg played an effective role in root induction in D. hamiltonii. Giridhar et al. [65] found that the shoots rooted on medium containing 9.8 μM IBA showed 80-90% survival rate under field conditions. While Gururaj et al. [66] found best rooting on 5.38 μM NAA and 400 μM PG containing MS medium. The efficient root formation was achieved when the elongated shoots of D. hamiltonii inoculated on medium supplemented with various phenolic compounds along with 9.8 μM IBA within 5-6 wk [65]. They found that PG and salicylic acid interaction with IBA stimulated maximum in vitro rooting of shoots [Table 2].
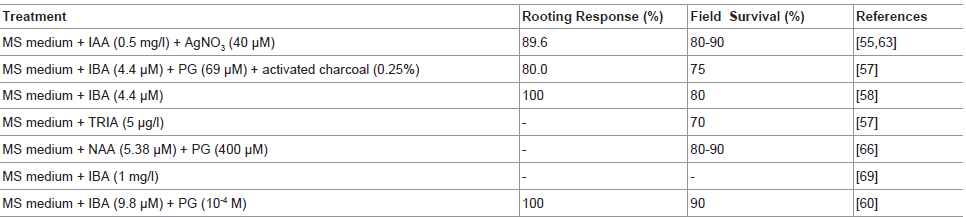
Table 2: Effect of different treatments on in vitro root induction of microshoots of D. hamiltonii (chronological order).
D. arayalpathra: There are only two reports for the micropropagation of D. arayalpathra in the year 2005. Gangaprasad et al. [59] cultured single node from young top shoots on MS medium supplemented with 0.1 to 5 mg/l BA. All the concentrations of BA induced single axillary shoots of varying length. However, MS medium supplemented with 0.1 mg/l BA supported rapid growth and produced the longest shoots (6.8 cm) in 60 days. For further multiplication, the nodes and shoot tips from in vitro derived shoots were re-cultured on MS medium fortified with 0.5 mg/l BA which produced 8 cm long shoot having 5-7 nodes in 30 days period. Further, the top microshoot cuttings (3-5 cm) with 2-3 nodes, subcultured onto MS medium supplemented with 1.5 mg/l IAA, produced an average of 6.3 roots in 30 days period. The rooted plants after hardening were reintroduced into their natural habitat at Kallar reserve forest, Thiruvananthapuram. After 2 years, 84% survival of reintroduced plants was recorded.
In another study of Sudha et al. [54] it was suggested that nodal explants of greenhouse-raised plants were more desirable than cotyledonary nodal explants of aseptic seedlings of D. arayalpathra. The basal nodes (73%) of 12-16-week-old greenhouse-grown plants cultured in MS medium containing 12.96 μM BA, 2.48 μM 2-iP and 2.68 μM NAA formed 16-17 cm long unbranched robust solitary shoots in 8 weeks. Cotyledonary nodal explants cultured in the same medium showed multiple shoot formation and axillary branching. But the shoots were thin, fragile and not suitable for mass propagation. Single nodes of a solitary shoot subcultured on MS medium containing 2.22 μM BA and 0.24 μM 2-ip together produced 9.8 nodes from 18.0 cm long shoots within 5-6 weeks. The basal nodes of the shoots so formed were repeatedly subcultured to increase the stock of propagules while 2.5-3.0 cm terminal cuttings were used for rooting. The best root induction (68%) and survival (86%) was achieved on half-strength MS medium supplemented with 1.07 μM NAA. Fieldestablished plants showed uniform growth and phenotypic similarity to parental stock [54].
Somatic Embryogenesis: As compared to the other species of Decalepis, somatic embryogenesis is only reported in D. hamiltonii through leaf culture by Giridhar et al. [69]. Nodular embryogenic callus was developed from the cut end of the explants on MS medium containing 2, 4-D and BA. On subsequent transfer of explants with primary callus on to the MS medium containing zea (13.68 μM) and BA (10.65 μM) resulted in the differentiation of somatic embryos directly from nodular tissue. The maturation of embryos took place on the same medium. The efficiency of somatic embryogenesis may enhanced when the embyrogenic calli with somatic embryos were subcultured onto MS basal medium supplemented with 4.56 μM zea and 10.65 μM BA. The mature embryos developed into complete plantlets on growth regulator-free MS medium.
Hydrogel Encapsulation: Encapsulation technology represents a new and powerful tool in the plant nursery field as well as in approaches to germplasm conservation and exchange of plant materials between laboratories [70]. Previously, synthetic seed or artificial seed (or synseed), described as ‘‘artificially encapsulated somatic embryos (bipolar structure) which can be used for sowing under in vitro or ex vitro conditions’’. But due to low success and high cost of somatic embryo production, shoot tips, nodes, bulbs or other meristematic tissue (unipolar structures) that can produce a whole plant may also be encapsulated which are also considered as synthetic seeds [71]. A wide range of woody plants has been re-grown from encapsulated shoot tips or nodes [72,73].
An efficient protocol has also been developed for short-term storage and conservation of D. hamiltonii using encapsulated nodal segments [74]. Only juvenile nodal segments, excised from aseptic seedlings were used for encapsulation. It was found that the encapsulation significantly affected by the concentrations of sodium alginate (Na-alginate) and calcium chloride (CaCl2·2H2O). A gelling matrix of 4% Na-alginate and 100 mM CaCl2·2H2O was most suitable for the production of ideal Ca-alginate beads. Maximum shoot regrowth (77.00%) was recorded on MS basal medium supplemented with 5.0 μM BA, 0.5 μM IAA and 30.0 μM adenine-sulphate (ADS) after 6 weeks of culture. Microshoots, recovered from encapsulated nodal segments (capsule) were best rooted on half-strength MS medium containing 2.5 μM NAA. Capsules of D. hamiltonii could be stored up to 8 weeks at low temperature (4°C) [75].
Being small in size the capsule, therefore can provide an effective tool for storage and exchange of this endemic and endangered plant species, potentially overcoming many of the difficulties associated with long-distance transport of plant germplasm. However, in this protocol rooting could not be possible in a single step on re-growth media which calls for further refinement in the established protocolin such a way that rooting could be achieved in a single step (on regrowth-medium).
Similar to the somatic embryogenesis, exploitation of encapsulation technique for the conservation of other species of Decalepis is still needed for their successful reintroduction in natural habitat.
Normal root culture
D. hamiltonii: Giridhar et al. [60] studied the production of 2H4MB by normal root cultures of D. hamiltonii. The flavour compound was extracted with dichloromethane, evaporated to dryness and dissolved in ethanol for qualitative (TLC) and quantitative (GCMS) analysis. Maximum root biomass and the maximum content of flavour compound (40 μgg-1 dry weight) were recorded after 45 days of growth on Murashige and Skoog (MS) medium containing 1.0 mgL-1α-naphthaleneacetic acid (NAA).
D. arayalpathra: Sudha and Seeni [35] reported for fast-growing normal root culture of D. arayalpathra from leaf and inter nodal explants of in vitro-raised shoot cultures in MS medium containing 2.5 mg/l BA, 0.5 mg/l 2-iP and 0.5 mg/l NAA. Shoot cultures were maintained on MS agar medium supplemented with 0.5 mg/l BA and 0.05 mg/l 2-iP or and 0.05 mg/l NAA and sub-cultured at 5 weeks interval. Leaf explants incubated in total darkness on half-strength MS medium supplemented with 0.5 mg/l IBA and 0.2 mg/l NAA favoured induction of roots (5.820 g). A 100 mg fresh root tissue cultured in 80 ml half-strength MS liquid medium supplemented with 0.2 mg/l IBA and 0.1 mg/l NAA, under continuous agitation (80 rpm), yielded 3.433 g fresh and 0.734 g dry weight of roots. Roots grown in this optimal medium produced maximum content of 2H4MB (0.16%) after 6 weeks of culture. The root cultures were maintained up to the 7th passage without decline of growth [35].
Hairy root culture
The normal root culture system was slow growing with a short life-span, as mentioned in an extensive review [76]. Advances in plant cell and tissue culture, combined with improvement in genetic engineering, specifically transformation technology has opened new avenues for high volume production of pharmaceuticals, nutraceuticals, and other beneficial substances [77]. Transgenic hairy root cultures have revolutionized the role of plant tissue culture in secondary metabolite production from the wild and commercial medicinal plants. Hairy root culture is an attractive experimental system, as they are long-term aseptic root clones, genetically stable with growth are comparable to those of the fastest-growing cell suspension culture [75]. This approach has been successfully applied for D. hamiltonii and D. arayalpathra as below;
D. hamiltonii: Samydurai et al. [78] reported Agrobacterium rhizogenes mediated hairy root culture and genetic transformation of D. hamiltonii. They have reported the total dry matter content of 2H4MB by hairy root culture of D. hamiltonii from internode and roots explants inoculated with A. rhizogenes. The addition of acetosyringone to basal MS liquid medium fortified with NAA and IBA considerably enhanced the hairy root production. PCR analysis of Ri plasmid on transformed internode and root tissues showed DNA amplification at 780 bp amplified in internode, root derived hairy roots. The highest biomass of dry matter content (25.36 and 24.12 g) was observed in liquid MS medium containing 0.3 mg/L NAA, cocultivated with A. rhizogenes on the internode and root explants. Afterthat the internode and root explants were cultured on MS medium supplemented with 0.3 mg/L IBA thereby exhibited improved biomass content of dry matter 23.56 and 22.91 g respectively. The dry matter content of biomass was extracted with 1 mg/ml methanol to measure the total content accumulation of 2H4MB in hairy roots cultures by using HPLC [77].
D. arayalpathra: Sudha et al. [79] studied A. rhizogenes mediated transformation of D. arayalpathra by infecting juvenile hypocotyl and cotyledon explants with different strains, including A4, MTCC 532, TR105 and LBA 5402. Hypocotyl explants induced hairy roots at a higher frequency (53.2 ± 0.3 %) than cotyledons (32.1 ± 0.2 %) when infected with the most virulent strain TR105. The explants co-cultivated 48 h in half-strength salts and vitamins of MS basal medium (half-MS medium) induced hairy roots either directly from the wounds or follwed by the formation of gall like structures. Irrespective of the explants, the strain MTCC 532 induced callus alone. The root initials on the galls proliferated vigorously and elongated more rapidly when they were segmented and subcultured on half-MS medium than the proliferation and elongation of directly emerged roots. The established hairy roots showed intermittent gall formation which was the active sites for hairy roots induction. The molecular evidence of rol A and rol C gene integration was confirmed by PCR amplification and southern blot hybridization. Growth of the hairy roots was undertaken by measuring root growth unit after culturing root tips in half-MS medium and determined fresh weight/dry weight/conductivity during time-course study in shake flask cultures. The maximum biomass and accumulation of the root specific compound, 2H4MB (0.22 % dry weight), was recorded at the 6th week of growth, which was more than that observed in normal root cultures (0.16 % dry weight) [78].
Conclusion
Decalepis is one of the most important woody medicinal climbing genera of the family “Periploaceace”. Until 1997 it remains the monotypic genera of this family, but now there are five species of Decalepis viz., D. hamiltonii, D. arayalpathra, D. salicifolia, D. khasiana and D. nervosa which are endemic to Eastern and Western Ghats of India. Amongst, D. hamiltonii is the most widespread and most utilized species. Except D. nervosa, roots are highly medicinal due to the presence of vallinin-like fragrant compound namely, 2H4MB. Ruthless collection of the roots is the main reason for the depletion of these species in nature and urgently calls for their conservation in natural habitats. Significant works have been carried out for the conservation of D. hamiltonii through plant tissue culture. While, only four reports are available for the D. araylpathra using plant tissue culture. So, there is a need to exploit all the possible valuable strategies of plant tissue culture like micropropagation, synseed preparation, genetic transformation and elicitation for other species of Decalepis. It is expected that the present article would provide an informative baseline to the researchers working on Decalepis species.
Acknowledgement
Dr. Shiwali Sharma is thankful to DST, for the award of Young Scientist under Fast Track Scheme, SERB (vide no. SB/FT/LS- 364/2012) for providing research assistance. Dr. Anwar Shahzad gratefully acknowledges the financial support provided by UGC and UP-CST in the form of research projects (vide no. 39-369/2010 SRand vide no. CST/D3836).
References
- Bapat VA, Yadav SR, Dixit GB (2008) Rescue of endangered plants through biotechnological applications. Nat Aca Sci Lett 31: 201-210.
- Phartyal SS, Thapliyal RC, Koedam N, Godefroid S (2002) Ex situ conservation of rare and valuable forest tree species through seed gene bank. Curr Sci 83: 1351-1357.
- Hamilton AC (2004) Medicinal plants, conservation and livelihoods. Biodiv Cons 13: 1477-1517.
- Ionta GM (2009) Phylogeny reconstruction of periplocoideae (apocynaceae) based on morphological and molecular characters and a taxonomic revision of Decalepis. The Ph.D. Thesis, University of Florida.
- Anonymous (2003) The Wealth of India: A Dictionary of Indian Raw Materials and Industrial Products. CSIR, New Delhi.Vol 3: 24.
- Giridhar P, Gururaj B and Ravishankar GA (2005) In vitro shoot multiplication through shoot tip cultures of Decalepis hamiltonii Wight & Arn., a threatened plant endemic to southern India. In Vitro Cell Dev Biol Plant 41: 77-80.
- Nayar MP (1989) In situ conservation of wild flora resources. Bull Bot Surv India 29: 319-333.
- Nayar MP (1996) Hot spots of endemic plants of India, Nepal and Bhutan. Tropical Botanical Garden Publishers, Thiruvananthapuram.
- Subramanyan K, Nayar MP (1974) Vegetation and phytogeography of the Western Ghats. Ecology and Biogeography of India. 23:178-196.
- Thenint A (1936) Un nouveau genre d’Apocynacées. Bull Soc Bot France 83: 389-393.
- Noltie HJ (2005) The botany of Robert Wight (Regnum vegetabile 145). Gartner Verlag, Liechtenstein.
- Venter HJT and Verhoeven RL (1997) A tribal classification of the Periplocoideae (Apocynaceae). Taxon 46: 705-720.
- Venter HJT, Verhoeven RL (2001) Diversity and relationships within the Periplocoideae (Apocynaceae). Ann Missouri Bot Gard 88: 550-568.
- Ionta GM and Judd WS (2007) Phylogenetic relationships in Periplocoideae (Apocynaceae s.l.) and insights into the origin of pollinia. Ann Missouri Bot Gard 94: 360-375.
- Meve U, Liede S (2001) Reconsideration of the status of Lavrania, Larryleachia and Notechidnopsis (Asclepiadoideae-Ceropegieae). South African J Bot 67: 161-168.
- Holmgren PK, Holmgren NH and Barnett LC (1990) Index Herbarium. Part I: The Herbaria of the World. New York Botanical Garden Press, New York.
- Vedavathy S (2004) Decalepis hamiltonii Wight & Arn.-an endangered source of indigenous health drink. Nat Product Rad 3: 22-23.
- Raju AJS, Ramana KV (2009) Pollination and seedling ecology of Decalepis hamiltonii Wight & Arn. (Periplocaceae), a commercially important endemic and endangered species. J Threatened Taxa 1: 497-506.
- Anandalakshmi R, Prakash MS (2009). Seed germination and storage characteristics of Decalepis hamiltonii implications for regeneration. Forest Tree Livelihood 19: 399-407.
- Saini RK, Giridhar P (2012) Effect of temperature, chemical treatment, hydration and plant growth regulators on germination of seeds and immature zygotic embryo of swallow root (Decalepis hamiltonii). Research J Agri Sci 3: 40-44.
- Murti PB, Seshadri TR (1941) A study of the chemical components of the roots of Decalepis hamiltonii (Makali veru), Part I- Chemical composition of the root. Proc Ind Acad Sci A 13: 221-232.
- Murti PB and Seshadri TR (1941) A study of the chemical components of the roots of Decalepis hamiltonii (Makali veru), Part II- A note on the preparation of inositol by solvent extraction. Proc Ind Acad Sci A 13: 263-265.
- Nagarajan SL, Rao LJM, Gurudatta KN (2001) Chemical composition of the volatiles of Decalepis hamiltonii Wight & Arn. Flavour Frag J 16: 27-29.
- George J, Pereira J, Divakar S, Udaysankar K and Ravishankar GA (1998) A method for preparation of active fraction from the root of Decalepis hamiltonii, useful as bioinsecticides. Indian patent no. 1301/Del/98.
- Thangadurai D, Anitha S, Pullaiah T, Reddy PN, Ramachandraiah S (2002) Essential oil constituents and in vitro antimicrobial activity of Decalepis hamiltonii roots against food borne pathogens. J Agric Food Chem 50: 3147-3149.
- Zhang Q, Zhao Y, Wang B, Feng R, Liu X, et al. (2002) New pregnane glycosides from Stelmatocrypton khasianum. Steroids 67: 347-351.
- Lakshman K, Jayaprakash B, Joshi H (2005) Comparative anti inflammatory activity studies of four species of sariva. Indian J Pharm Sci 67: 481-482..
- Nagarajan S, Rao LJM (2003) Determination of 2-hydroxy-4-methoxybenzaldehyde in roots of Decalepis hamiltonii (Wight & Arn.) and Hemidesmus indicus R. Br. J AOAC Int 86: 564-567.
- Srivastava A, Harish RS, Shivanandappa T (2006) Novel antioxidant compounds from the aqueous extract of the roots of Decalepis hamiltonii Wight & Arn. and their inhibitory effect on low-density lipoprotein oxidation. J Agri Food Chem 54: 790-795.
- Srikanta BM, Siddaraju MN, Dharmesh SM (2007) A novel phenol bound pectic polysaccharide from Decalepis hamiltonii with multi-step ulcer preventive activity. World J Gastroenterol 13: 5196-5207.
- Chopra RN, Nayar SL, Chopra LC, Asolkar LV, Kakkar KK (1956) Glossary of Indian medicinal Plants; Council of Scientific and Industrial Research, New Delhi.
- Pushpangadan P, Rajasekharan A, Ratheeshkumar PK, Jawahar CR, Radhakrishnan K, et al. (1990) Amrithapala (Janakia arayalpathra Joseph & Chandrasekharan), a new drug from the Kani tribe of Kerala. Anc Sci Life 9: 212-214.
- Radhakrishnan K, Pandurangan AG, Pushpangadan P (1998) Utleria salicifolia-a new ethnobotanical record from Kerala, India. Fitoterapia 69: 403-405.
- Thénint A (1936) Un nouveau genre d’Apocynacées. Bull Soc Bot France 83: 389-393.
- Sudha CG, Seeni S (2001) Establishment and analysis of fast-growing normal root culture of Decalepis arayalpathra, a rare endemic medicinal plant. Curr Sci 81: 371-374.
- Nayar RC, Shetty JKP, Mary Z, Yoganarshimhan SN (1978) Pharmacognostical studies on the root of Decalepis hamiltonii and comparison with Hemidesmus indicus. Proc Indian Acad Sci 87: 37-48.
- Ashalatha K, Venkateswarlu Y, Priya AM, Lalitha P, Krishnaveni M, et al. (2010) Anti-inflammatory potential of Decalepis hamiltonii (Wight and Arn.) as evidenced by down regulation of pro inflammatory cytokines-TNF-alpha and IL-2. J Ethnopharmacol 130: 167-170.
- Jacob KC (1937) An unrecorded economic product Decalepis hamiltonii W. & Arn., family Asclepidaceae. Madras Agric J 25: 176.
- Vijayakumar V and Pullaiah T (1998) An ethno-medico-botanical study of Prakasam district, Andhra Pradesh, India. Fitoterap 69: 483-489.
- Harish R, Divakar S, Srivastava A, Shivanandappa T (2005) Isolation of antioxidant compounds from the methanolic extract of the roots of Decalepis hamiltonii (Wight & Arn.). J Agric Food Chem 53: 7709-7714.
- Naveen S, Khanum F (2010) Antidiabetic, antiatherosclerotic and hepatoprotective properties of Decalepis hamiltonii in streptozotocin-induced diabetic rats. 34: 1231-1248.
- Joseph J, Chandrasekaran V (1978) Janakia arayalpathra-a new genus and species of Periplocaceae from Kerala, South India. J Indian Bot Soc 57: 308-312.
- Mohana DC, Raveesha KA (2007) Anti-fungal evaluation of some plant extracts against some plant pathogenic field and storage fungi. JAT 4: 119-137.
- Mohana DC, Satish S, Raveesha KA (2009) Antifungal activity of 2-hydroxy-4- methoxybenzaldehyde isolated from Decalepis hamiltonii Wight & Arn on seed-borne fungi causing biodeterioration of paddy. J Plant Prot Res 49: 250-256.
- Thangavel K, Ebbie MG, Ravichandaran P (2011) Antibacterial potential of Decalepis hamiltonii Wight & Arn. callus extract. Nat Pharma Tech 1: 14-18.
- Devi M and Latha P (2012) Antibacterial and phytochemical studies of various extracts of roots of Decalepis hamiltonii Wight and Arn. Int J Pharma Pharmaceutical Sci 4: 738-740.
- George J, Udaysankar K, Keshava N, Ravishankar GA (1999) Antibacterial activity of supercritical extract from Decalepis hamiltonii roots. Fitoterap 70: 172-174.
- Halliwell B, Gutteridge JM (1999) Free radicals in biology and medicine. Oxford: Oxford University Presss.
- Murthy KN, Rajasekaran T, Giridhar P, Ravishankar GA (2006) Antioxidant property of Decalepis hamiltonii Wight & Arn. Indian J Exp Biol 44: 832-837.
- Srivastava A, Rao JM, Shivanandappa T (2007) Isolation of ellagic acid from the aqueous extract of the roots of Decalepis hamiltonii: antioxidant activity and cytoprotective effect. Food Chem 103: 224-233.
- Kumuda KV, Shashidhara S, Rajasekharan PE, Ravish BS (2011) Study of in vitro anti typhoid activity of various roots extracts of Decalepis hamiltonii (Wight & Arn.). Int J Pharma Biol Archives 2: 546-548.
- Sumalatha G, VidyaSagar J, Ragini V, Suresh K (2010) Extraction and evaluation of roots of Decalepis hamiltonii for antidiabetic activity. Int J Pharmacognosy Phytochem Res 2: 20-25.
- Zarei M, Shivanandappa T (2013) Amelioration of cyclophosphamide induced hepatotoxicity by the root extract of Decalepis hamiltonii in mice. Food Chem Toxicol 57: 179-184.
- CG, Krishnan PN, Pushpangadan P, Seeni S (2005) In vitro propagation of Decalepis arayalpathra, a critically endangered ethnomedicinal plant In Vitro Cell Dev Biol- Plant 41: 648-654.
- Bais HP, George J and Ravishankar GA (2000) In vitro propagation of Decalepis hamiltonii Wight & Arn. An endangered shrub through axillary bud cultures. Curr Sci 79: 408-410.
- Raju AJS (2010) Pollination Biology of Decalepis hamiltonii and Shorea tumbuggai Lambert Academic Publishing 64.
- Reddy OB, Giridhar P, Ravishankar GA (2002) The effect of triacontanol on micropropagation of Capsicum frutescens and Decalepis hamiltonii W & A. Plant Cell Tiss Org Cult 71: 253-258.
- Reddy OB, Giridhar P, Ravishankar GA (2001) In vitro rooting of Decalepis hamiltonii Wight & Arn., an endangered shrub, by auxins and root promoting agents. Curr Sci 81: 1479-1482.
- Gangaprasad A, Decruse SW, Seeni S, Nair GM (2005) Micropropagation and ecorestoration of Decalepis arayalpathra (Joseph & Chandra.) Venter-an endemic and endangered ethnomedicinal plant of Western Ghats. Indian J Biotechnol 4: 265-270.
- Giridhar P, Rajasekaran T, Ravishankar GA (2005) Improvement of growth and root specific flavour compound 2-hydroxy-4-methoxy benzaldehyde of micropropagated plants of Decalepis hamiltonii Wight & Arn., under triacontanol treatment. Sci Hort 106: 228-236.
- Giridhar P, Rajasekaran Y, Ravishankar GA (2005) Production of root specific flavour compound, 2-hydroxy-4-methoxy benzaldehyde by normal root cultures of Decalepis hamiltonii Wight & Arn (Asclepiadaceae). J Sci Food Agri 85: 61-64.
- George J, Bais HP and Ravishankar GA (2000) Optimization of media constituents for shoot regeneration from leaf callus cultures of Decalepis hamiltonii Wight & Arn. Hort Sci 35: 296-299.
- Bais HP, Sidha G, Suresh B, Ravishankar GA (2000) AgNO3 influences in vitro root formation in Decalepis hamiltonii Wight & Arn. Curr Sci 79: 894-898.
- Anitha S, mPullaiah T (2002) In vitro Propagation of Decalepis hamiltonii. J Trop Med Plants 3: 227-232.
- Giridhar P, Ramu DV, Reddy BO, Rajasekaran T, Ravishankar GA (2003) Influence of phenylacetic acid on clonal propagation of Decalepis hamiltonii Wight & Arn an endangered shrub. In Vitro Cell Dev Biol-Plant 39: 463-467.
- Gururaj HB, Giridhar P, Ravishankar GA (2004) Efficient clonal propagation method for Decalepis hamiltonii an endangered shrub, under the influence of phloroglucinol. Indian J Exp Biol 42: 424-428.
- Giridhar P, Gururaj B and Ravishankar GA (2005) In vitro shoot multiplication through shoot tip cultures of Decalepis hamiltonii Wight & Arn., a threatened plant endemic to southern India. In Vitro Cell Dev Biol Plant 41: 77-80.
- Giridhar P, Rajasekaran T, Nagarajan S, Ravishankar GA (2004) Production of 2-hydroxy-4-methoxy benzaldehyde in roots of tissue culture raised and acclimatized plants of Decalepis hamiltonii Wight & Arn an endangered shrub endemic to Southern India and evaluation of its performance vis-Ã -vis plants from natural habitat. Indian J Exp Biol 42: 106-110.
- Giridhar P, Kumar V, Ravishankar GA (2004) Somatic embryogenesis, organogenesis and regeneration from leaf callus culture of Decalepis hamiltonii Wight & Arn., an endangered shrub. In Vitro Cell Dev Biol-Plant 40: 567-571.
- Sharma S, Shahzad A, Teixeira da Silva JA (2013) Synseed technology-a complete synthesis. Biotechnol Adv 31: 186-207.
- Pond S and Cameron S (2003). Tissue culture: artificial seeds. In: Thomas B, Murphy DJ, Murray BG (Eds.) Encyclopaedia of Applied Plant Sciences. Amsterdam: Elsevier Academic Press 1379-1388.
- Chand S, Singh AK (2004) Plant regeneration from encapsulated nodal segments of Dalbergia sissoo Roxb.-a timber yielding leguminous tree. J Plant Physiol 161: 237-243.
- Rai MK, Asthana P, Singh SK, Jaiswal VS , Jaiswal U (2009) The encapsulation technology in fruit plants-a review. Biotechnol Adv 27: 671-679.
- Germanà MA, Micheli M, Chiancone B, Macaluso L, Standardi A (2011) Organogenesis and encapsulation of in vitro-derived propagules of Carrizo citrange (Citrus sinensis (L.) Osb. × Poncirius trifoliate (L.) Raf.). Plant Cell Tiss Organ Cult 106: 299-307.
- Sharma S, Shahzad A (2012) Encapsulation technology for short-term storage and conservation of a woody climber, Decalepis hamiltonii Wight and Arn. Plant Cell Tiss Org Cult 111: 191-198..
- Giri A and Narasu ML (2000) Transgenic hairy roots: recent trends and applications. Biotechnol Advan 18: 1-22.
- Hansen G and Wright MS (1999) Recent advances in the transformation of plants. Trends Plant Sci 4: 226-231.
- Samydurai P, Ramakrishnan R, Thangapandian V (2013) Agrobacterium rhizogenes mediated hairy root culture and genetic transformation of an endangered medicinal plant of Decalepis hamiltonii Wight & Arn. J Microbiol Biotechnol Food Sci 3: 191-194.
- Sudha CG, Sherina TV, Anu Anand VP, Reji JV, Padmesh P, Soniya EV (2013) Agrobacterium rhizogenes mediated transformation of the medicinal plant Decalepis arayalpathra and production of 2-hydroxy-4-methoxy benzaldehyde. Plant Cell Tiss Org Cult 112: 217-226.