Research Article
Fission Yeast Topoisomerase II (Top2) isRequired for the Viability of Pot1Δ Single Mutant
Ahmed G. G. Darwish1,2*, Mamdouh N. Samy2,3, Hosni Abdel-Salam1 and Emad E. Shaker1
Corresponding author: Dr. Ahmed G. G. Darwish, a Department of Biochemistry, Faculty of Agriculture, Minia University,Minia 61519, Egypt,; E-mail: gomaaahmed75@yahoo.com
Citation: Darwish AGG, Samy MN, Abdel-Salam H, Shaker EE. Fission Yeast Topoisomerase II (Top2) is Required for the Viability of Pot1Δ Single02 Mutant. J Cell Sci Molecul Biol. 2015;2(1): 113.
Copyright © 2015 Ahmed G. G. Darwish et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Submission: 08/04/2015; Accepted: 09/05/2015; Published: 15/05/2015
Abstract
Topoisomerase II constitutes a group of nuclear enzymes which alters the topological state of DNA. These enzymes have the ability to transferring one DNA double helix through a transient break in another DNA double helix. Topoisomerases type II have important roles in DNA metabolic processes such as DNA replication, transcription and chromosome segregation, condensation and de-condensation. Recently Topoisomerases II become very important target for many anticancer drugs such as Doxorubicin, Etoposide, and Aminoacridines. So, our results suggest that Topoisomerase II is required for the viability of pot1Δ cells by creation fission yeast (Shizosaccharomyces Pombe) double mutant model (pot1Δ top2Δ). In addition to the deletion of Rav1 gene enhance the sensitivity of cells to drugs. This leads to developing assay system to screening Topoisomerase II inhibitors by creation (S. Pombe) double mutant model pot1Δ rav1Δ.
Keywords: DNA; Doxorubicin; Schizosaccharomyces Pombe; pot1; top2; rav1.
Introduction
Schizosaccharomyces Pombe is a single celled free livingarchiascomycete fungus sharing many features with cells is morecomplicated eukaryotes. From gene sequence comparisons andphylogenetic analyses it has been suggested that fission yeast divergedfrom budding yeast around 330-420 million years ago, and frommetazoan and plants around 1,000-1,200 million years ago [1]. Somegene sequences are as equally diverged between the two yeasts asthey are from their human homologues, probably reflecting a morerapid evolution within fungal lineages than in the metazoan S. pombewas first described in the 1890s and has been extensively studiedsince the 1950s [2,3]. It has served as an excellent model organismfor the study of cell cycle control, mitosis [4], DNA repair andrecombination [5] and checkpoint controls important for genomestability [6]. The 13.8 Mb genome of S. pombe is distributed between chromosomes I (5.7 Mb), II (4.6 Mb) and III (3.5 Mb) [7], togetherwith a 20 Kb mitochondrial genome [8]. Topoisomerase II is locatedat chromosome II (1.976 Mb 1.980 Mb).
DNA Topoisomerases type 2 is essential enzymes to all living cells.These enzymes have the ability to transferring one DNA double helixthrough a transient break in another DNA double helix [8], whileTopoisomerases type 1 makes single strand breaks through the nickto allow relaxation of over-coiled DNA [9]. In eukaryotes the enzymescatalyzes the ATP- dependent relaxation of negatively and positivelysupercoiled DNA as well as catenation-decatenation and knottingunknottingcircular DNA [10,11] and have important roles in DNAreplication, transcription and chromosome segregation, chromosomecondensation and decondensation. Top2 gene is also very importanttarget for a widely number of anticancer drugs in clinical use, such asthe anthracyclines [12], which acts as inhibitor for the DNA topology regulating enzyme top2. Inhibiting of top2 results in accumulationof DSB that leads to cell death [13]. Furthermore, Topoisomerasepoisons can be categorized into three groups; intercalators (such asdoxorubicin and m-AMSA), non-DNA-intercalators (etoposide,teniposide) and catalytic inhibitors of the topoisomerase (aclarubicinand bisdioxopiperidines like ICRF-193) [14]. The first two groupstrap top2 on the DNA in a covalently bound state, often referred to asthe cleavable complex [15]. Catalytic inhibitors act on top2 withoutforming the cleavable complex. It is not entirely clear what causes celldeath following topoisomerase stabilization. DNA damage provokedby top2 poisons was shown in some cases to result in apoptosis [16].
Mechanism of drug resistance
Drug resistant topoisomerase 2 restricts the therapeutic choicesin many cases. It is sometimes caused by the overexpression ofmembrane P-glycoprotein (MDRl gene product) which responsibleof efflux for many drugs from cells [17]. Others are produced by theMDRl independent mechanisms, which in many cases are correlatedwith alterations of top2 gene, like the expression level or mutation.The expression level of top2 is affected by several factors, especially,down regulation of the top2 mRNA [18], instability of mRNA [19]and destruction of the top2 protein [20].
Identification of top2 inhibitors resistance genes in fissionyeast
Protection of telomeres protein 1 (pot1) binds to single- stranded telomere overhangs and protects chromosome ends [21]. In S. pombe the deletion of pot1 causes rapid telomere loss and chromosomecircularization [22] by single strand annealing (SSA), suggesting thatthe chromosome ends are resected in the absence of pot1 [23,24] gene.
Rav1 gene encodes the homologue of a budding yeast proteinwhich regulates the assembly of vacuolar ATPase this gene haveimportant role in Multi drug resistance (MDR) [25,26]. Themutation in this gene (rav1Δ) make the cell very sensitive to thechemotherapeutic agents .We found that rav1Δ mutant leads toincrease the accumulation of the drug in cells [27]. The function ofRav1 appears to be conserved, S. Pombe rav1 is part of a RAVE-likecomplex in fission yeast and loss of rav1 gene results in an increasedsensitivity of S. pombe cells to drugs [28].
The aim of our work was investigate that top2 gene is required for the viability of pot1Δ cells using S. Pombe pot1Δ top2Δ double mutant and developing assay system to screening top2 inhibitors using S. Pombe double mutant pot1Δ rav1Δ as a sensitive model to drugs.
Materials and Methods
Strain construction and growth media
Fission yeast 972 (h-) strain, YEA (3% glucose, 0.5% yeast extract,and 75 mg/L adenine) media was used to grow fission yeast cells. pot1Δrav1Δ double mutant (pot1::KanMX rav1::hphMX) expressing pot1from a plasmid (ppc27-pot1-hemaagglutinin [HA]) was created bygenetic interaction (mating cells) between pot1::kanMX Δ (h+) singlemutant which have pot1 plasmid with rav1::hphMX Δ single mutant(h-) using ME medium (minimal medium Bacto-malt extract 30 g/L)adjusting PH 5.5. The double mutant were selected on Kan plate (YEA containing Kanamycin 1 μL/ mL) , hph plate (YEA containingHygromycin 1 μL/ mL) and yeast extract agar (YEA) plates containing2 g/ L 5-Fluoroorotic acid (FOA) and EMM3+ALH plate (Edinburghminimal media containing alanine, leucine and histidine). The loss oftelomeric repeats and chromosome circularization were confirmed bysouthern hybridization and pulsed-field gel electrophoresis (PFGE)respectively.
pot1Δ top2Δ double mutant (pot1::KanMX top2:: ura4) expressing Pot1 from a plasmid (ppc27-pot1-hemaagglutinin [HA], containing the ura4 gene) was created by genetic interaction (mating cells) between pot1::KanMX Δ (h+) single mutant which have pot1 plasmidwith top2::ura4Δ (h-) single mutant using ME medium, the doublemutant were selected on yeast extract agar (YEA) plates containing 2g/ L 5-Fluoroorotic acid (FOA).
Chemicals and solvents
Kanamycin (sigma), hygromycin (sigma), yeast extract (Becton,Dickinson. Company), AlKphos direct kit module (GE Healthcare),EcoRI restriction enzyme and NotI enzyme (Bio labs. Inc.), otherchemicals described later.
Measurement of Telomere Length telomere length was measuredusing southern hybridization with an AlKPhos Direct Kit module(GE Healthcare), according to a previously described procedure [29].
Pulsed-Field Gel Electrophoresis (PFGE) was performed asdescribed by Baumann et al. [30] for The detection of Not I-digestedchromosomes, Not I-digested S.pombe chromosomal DNA wasfractionated in a 1% agarose gel with 0.5×TBE (50 mM Tris-HCl, 5mM boric acid and 1 mM EDTA, pH 8.0) buffer at 14 °C using theCHEF Mapper PFGE system at 6 v/m (200 v) and a pulse time of60 to 120 s for 24 h. DNA was visualized by staining with ethidiumbromide (1μg/ ml) for 30 min.
Western blot analysis Cells were lysed in 10% trichloroaceticacid (TCA) with glass beads at 4 °C. After centrifugation at 15,000× g for 10 min at 4 °C, the precipitate was washed with acetone andsuspended in SDS sample buffer.
Microscopy Microscope images of living cells were obtainedusing an AxioCam digital camera (Zeiss) connected to an Axioobserver Z1 micro-scope (Zeiss).
DAPI stained a 10×DAPI mounting stock prepared andaliquoted. This is 10 μg/ml DAPI, 10 mg/ml p-phenyl enediamine.(The DAPI stock is 1 mg/ml in DMSO) take a 20 μl aliquot and add180 μl glycerol this gives a 1×working stock (1 μg/ ml DAPI, 1 mg/ml p-phenylenediamine which acts as anti-fade, 90% glycerol) whichshould be kept at -20 °C in the dark.
Cell counting with a Hemocytometer 10 μl cell culture in YEAliquid medium and the cell visualized by normal microscope after 4,16, 32, 64, 128 hours.
Flow cytometry
Cell Staining With PI
Suspend approx 106cells in 0.5 mL of PBS. Vortex gently (approx5 s) to obtain a mono-dispersed cell suspension, with minimal cell aggregation ,then fix cells by transferring this suspension witha Pasteur pipet into centrifuge tubes containing 4.5 mL of 70%ethanol on ice. Keep cells in ethanol for at least 2 h at 4 °C, after thatCentrifuge the ethanol-suspended cells for 5 min at 300 g, Suspendthe cell pellet in 5 mL of PBS wait approx 30 s and centrifuge at 300 gfor 5 min, then Suspend the cell pellet in 1 mL of PI staining solution.Keep in the dark at room temperature for 30 min, or at 37 °C for 10min, finally Transfer sample to the flow cytometer and measure cellfluorescence maximum excitation of PI bound to DNA is at 536 nm,and emission is at 617 nm blue (488nm).
Fluorescence Activated Cell Sorting (FACS) analysis usingdesktop flow cytometer the Becton-Dickinson fluorescence activatedcell sorter.
Spotting assay for growth
Strains were tested by growing to exponential phase followedby spotting individual ten-fold serially diluted strains manually onagar media containing doxorubicin (DOXO), hydroxyl urea (HU),thiabendazole (TBZ) and methyl methane sulphonate (MMS) atdifferent concentrations and incubated at 30 °C for three days or fourdays as follow, grow strains overnight in 5 ml liquid medium (e.g.YEA) at 30 °C, then Measure OD600and prepare dilutions of OD600=1 with sterile H2O, dilution series: 100, 10-1, 10-2, -3 (use spottingplate), then drop dilutions onto appropriate plates using a sterilemetal stamper, creating a grid of descending cell concentrations. Letplates dry completely, Incubate at 30 °C. Finally, take photographsat different days of incubation since differences can change quitedrastically during growth.
Results
The phenotypic of the pot1Δ top2Δ double mutant our resultshowed that after we streaked pot1Δ top2Δ double mutant, Wild type(W.T), Pot1Δ single mutant which lost for pot1 plasmid and top2Δsingle mutant on YEA medium plate and incubated at differenttemperatures 25 °C, 30 °C. The result shows pot1Δ top2Δ is notviable at 30 °C, but pot1Δ and top2Δ is viable compare to W.T as acontrol Figure 1. Our results suggest that top2 gene function requiredto maintain viability under these conditions. This might occur afterthe circularization and suggested that top2 gene efficiency verylow and disorders happened in chromosome segregation or DNAsynthesis. If this were the case we must detect the problems duringS phase by performed FACS analysis to detect the problems duringDNA synthesis in addition to detect problems during M phase byperformed DAPI stained.
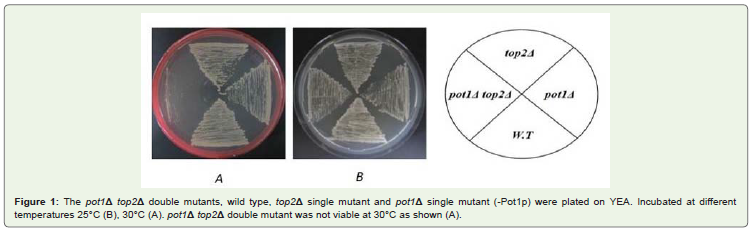
Figure 1: The pot1Δ top2Δ double mutants, wild type, top2Δ single mutant and pot1Δ single mutant (-Pot1p) were plated on YEA. Incubated at differenttemperatures 25°C (B), 30°C (A). pot1Δ top2Δ double mutant was not viable at 30°C as shown (A).
FACS analysis
The analysis of cellular DNA content following cell staining withboth propidium Iodide (PI) and deconvolution of the cellular DNAcontent frequency histograms. This approach reveals distribution ofcells in three major phases of the cell cycle (G1, S, and G2/M). Thedata are presented as cellular DNA content frequency histograms. Toobtain percentage of cells in S phase to detect the disorders in DNAsynthesis during S- phase. Our results showed that the pot1Δ top2Δdouble mutant cells have deficiency in DNA synthesis compare topot1 single mutant and Top2 single mutant under these condition asshown in Figure 2-Figure 4.
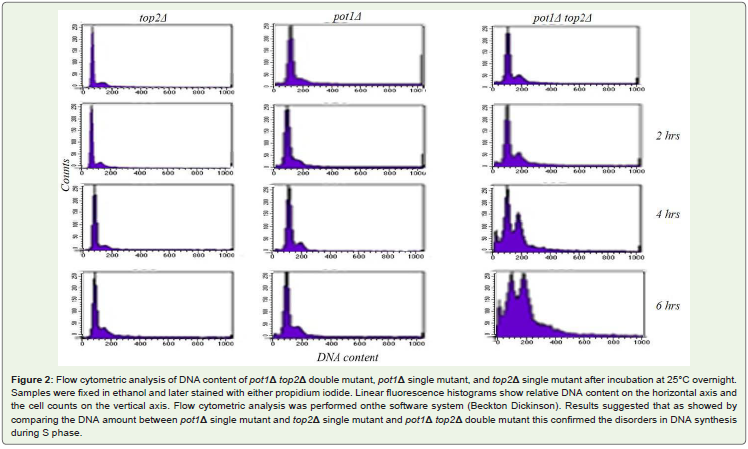
Figure 2: Flow cytometric analysis of DNA content of pot1Δ top2Δ double mutant, pot1Δ single mutant, and top2Δ single mutant after incubation at 25°C overnight.Samples were fixed in ethanol and later stained with either propidium iodide. Linear fluorescence histograms show relative DNA content on the horizontal axis andthe cell counts on the vertical axis. Flow cytometric analysis was performed onthe software system (Beckton Dickinson). Results suggested that as showed bycomparing the DNA amount between pot1Δ single mutant and top2Δ single mutant and pot1Δ top2Δ double mutant this confirmed the disorders in DNA synthesisduring S phase.
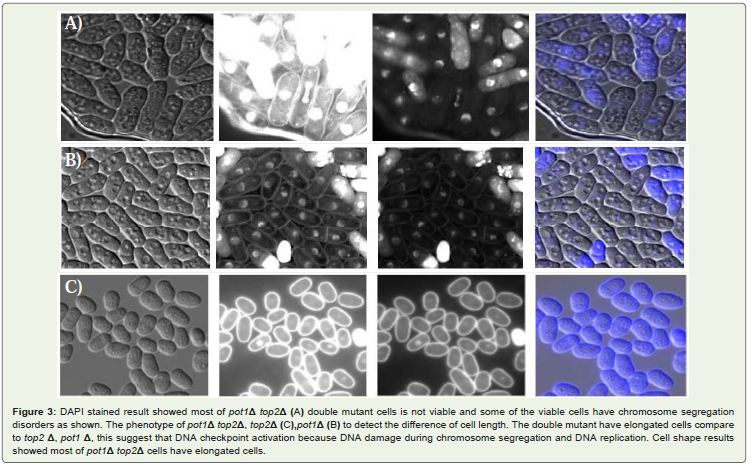
Figure 3: DAPI stained result showed most of pot1Δ top2Δ (A) double mutant cells is not viable and some of the viable cells have chromosome segregationdisorders as shown. The phenotype of pot1Δ top2Δ, top2Δ (C),pot1Δ (B) to detect the difference of cell length. The double mutant have elongated cells compareto top2 Δ, pot1 Δ, this suggest that DNA checkpoint activation because DNA damage during chromosome segregation and DNA replication. Cell shape resultsshowed most of pot1Δ top2Δ cells have elongated cells.
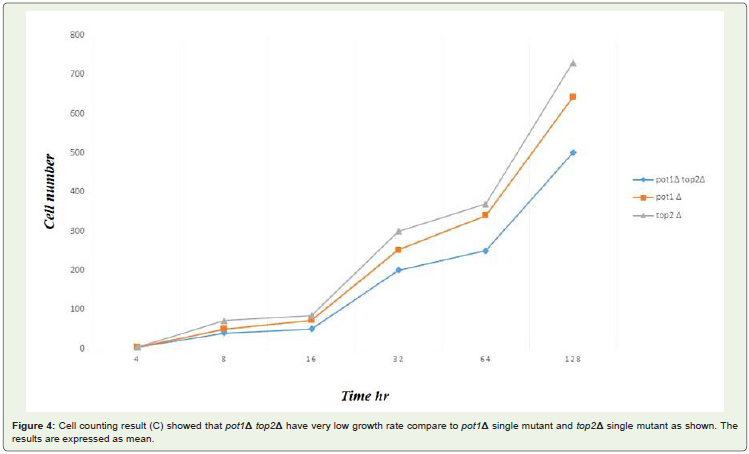
Figure 4: Cell counting result (C) showed that pot1Δ top2Δ have very low growth rate compare to pot1Δ single mutant and top2Δ single mutant as shown. Theresults are expressed as mean.
Both candidates of pot1Δ rav1Δ double mutant lose telomericDNA completely, and the chromosomes are circularized. The pot1Δdisruptant loses telomeric DNA completely, and survival depends onthe circularization of chromosome via SSA [31]. We analyzed telomerelength in the double mutants by using southern hybridization, bothcandidates of pot1Δ rav1Δ double mutant had lost telomeric DNAFigure 5. Next, we analyzed the chromosome structure by Pulsed FieldGel Electrophoresis (PFGE) Figure 6. The Not I-digested fragmentsM, L, I, and C, which are located in wild type cells. In contrast, the L+I band can be detected in both candidates of pot1Δ rav1Δ doublemutant. This is similar to pot1Δ single mutant, which has circularchromosomes. This results demonstrate that the chromosomes ofboth the pot1Δ rav1Δ double mutants are circularized.
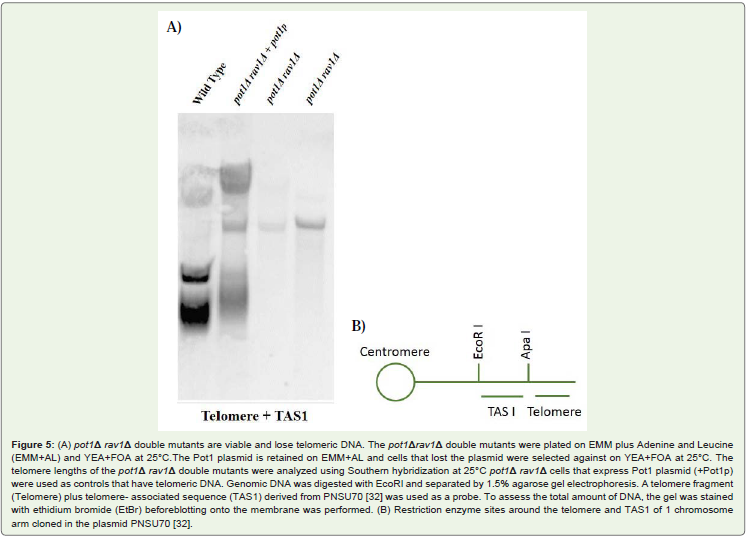
Figure 5: (A) pot1Δ rav1Δ double mutants are viable and lose telomeric DNA. The pot1Δrav1Δ double mutants were plated on EMM plus Adenine and Leucine(EMM+AL) and YEA+FOA at 25°C.The Pot1 plasmid is retained on EMM+AL and cells that lost the plasmid were selected against on YEA+FOA at 25°C. Thetelomere lengths of the pot1Δ rav1Δ double mutants were analyzed using Southern hybridization at 25°C pot1Δ rav1Δ cells that express Pot1 plasmid (+Pot1p)were used as controls that have telomeric DNA. Genomic DNA was digested with EcoRI and separated by 1.5% agarose gel electrophoresis. A telomere fragment(Telomere) plus telomere- associated sequence (TAS1) derived from PNSU70 [32] was used as a probe. To assess the total amount of DNA, the gel was stainedwith ethidium bromide (EtBr) beforeblotting onto the membrane was performed. (B) Restriction enzyme sites around the telomere and TAS1 of 1 chromosomearm cloned in the plasmid PNSU70 [32].
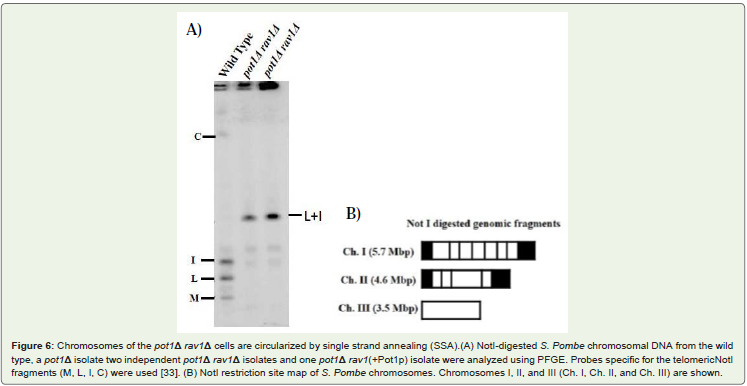
Figure 6: Chromosomes of the pot1Δ rav1Δ cells are circularized by single strand annealing (SSA).(A) NotI-digested S. Pombe chromosomal DNA from the wildtype, a pot1Δ isolate two independent pot1Δ rav1Δ isolates and one pot1Δ rav1(+Pot1p) isolate were analyzed using PFGE. Probes specific for the telomericNotIfragments (M, L, I, C) were used [33]. (B) NotI restriction site map of S. Pombe chromosomes. Chromosomes I, II, and III (Ch. I, Ch. II, and Ch. III) are shown.
Sensitivity of pot1Δ rav1Δ double mutant to drugs The viabilityof a mutant of the gene encoding a regulator of assembly of V-ATPase(rav1Δ) and Both pot1Δ rav1Δ candidates showed hypersensitivityto Doxorubicin as reported before [33] at final concentration(10 μg/ml) and hydroxyurea (HU) at final concentration (4 mM) and slightlyhigh sensitivity to hydroxyurea at final concentration (2 mM) andmoderate sensitivity to thiabendazole (TBZ) at final concentration (10mg/ml) and Miracle Mineral Solution (MMS) at final concentration(0.003% and 0.0015%) Figure 7.
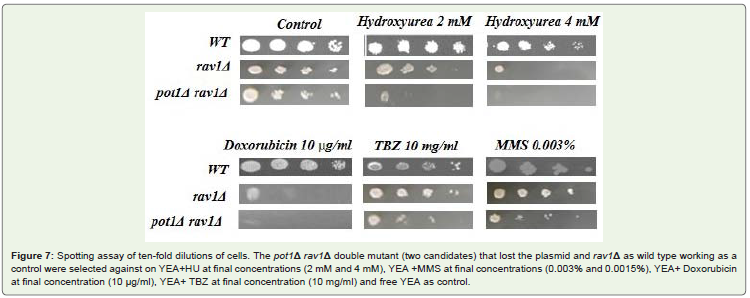
Figure 7: Spotting assay of ten-fold dilutions of cells. The pot1Δ rav1Δ double mutant (two candidates) that lost the plasmid and rav1Δ as wild type working as acontrol were selected against on YEA+HU at final concentrations (2 mM and 4 mM), YEA +MMS at final concentrations (0.003% and 0.0015%), YEA+ Doxorubicinat final concentration (10 μg/ml), YEA+ TBZ at final concentration (10 mg/ml) and free YEA as control.
Discussion
pot1Δ cells survival of the loss of telomere function is due tochromosome circularization. That is depended on SSA. It has beenrecently suggested that S. Pombe top2 gene has a similar involvement.This led us to investigate the hyposis that top2 gene required for theviability of pot1Δ cells Figure 1. FACS analysis results suggested thatthe unviability of pot1Δ top2Δ double mutant due to the disorders inDNA synthesis during S phase Figure 2. DAPI stained results gave usanother hyposis for the unviability of the double mutant might due tothe some problems happen during chromosome segregation duringM phase Figure 3. In addition to the growth of the double mutant at30°C gave us very clear evidence when the efficiency of top2 gene wasvery low the pot1Δ cells couldn’t grow.
top2 gene very important target for many new anticancer drugsthis led us to developing a new assay system using S. Pombe pot1Δrav1Δ double mutant to use these model for screening top2 inhibitors.The mutation in rav1 gene have increased the sensitivity of pot1Δcells toward drugs as shown in Figure 7. This evidence gave us theimportant role of rav1 gene in drug efflux and influx.
Acknowledgement
The authors are very grateful for access to FACS analysis, PFGE and Southern hybridization instruments at the Graduate School ofAdvanced Science of Matter and Graduate School of Biomedical andHealth Science Hiroshima University.
References
- Sipieki M (2000) Where does fission yeast sit on the tree of life?. Genome Biol 4: 1-1011.
- Leupold U (1950) Die Verebung Von Homothallic and Heterothallic bie Schizosaccharomyces Pombe. C R Lab Carlsberg 24: 381-475.
- Mitchison JM (1957) The growth of single cells: I. Schizosaccharomyces Pobme. Exp Cell Res 13: 244-262.
- Fantes P, Beggs J (2000) The yeast Nucleus (oxford Univ Press, Oxford, 2000).
- Daris L, Smith GR (2001) Meiotic recombination and chromosome segregation in Schizosaccharomyces Pombe. Proc Nat I Acad Sci USA 98: 8395-8402.
- Humphrey T (2000) DNA damage and Cell cycle control in Schizosaccharomyces Pombe. Mutant. Res, 451: 211-226.
- Lang BF, Cedergrn R, Gray MW (1987) The mitochondrial genome of the fission yeast, Schizosaccharomyces Pobme. Sequence of the large-subunit ribosomal RNA gene, comparison of potential secondary structure in fungal mitochondrial large- Subunit rRNAs and evolutionary considerations. Eur J Biochem 169: 527-537.
- Devendra ST (2011) Topoisomerase II inhibitors in cancer treatment. International Journal of Pharmaceutical Sciences and Nanotechnology: 3.
- Kellner U, Sehested M, Jensen PB, Gieseler F, Rudolph P (2002) Culprit and Victim- DNA topoisomerase II, Lancet Oncol 3: 235-243.
- Hsieh TS, Brutlag D (1980) ATP-dependent DNA topoisomerase from D. melanogaster reversibly catenates duplex DNA rings. Cell 21: 115-125.
- Baldi M, Beneditti P, Mattoccia, E, Tocchini-ValentiniGP(1980) In vitro catenation and decatenation of DNA and a novel eukaryotic ATP-dependent topoisomerase. Cell 20: 461-467.
- Hartsuiker E, Bahler J, Kohli J (1998) The role of Topoisomerase II in meiotic chromosome Condensation and segregation in Schizosaccharomyces Pombe. Mol Biol Cell 9: 2739-2750.
- Pommier Y, Leo E, Zhang H, Marchand C (2010) DNA topoisomerases and SWI/SNF and RSC complexes show compositional and functional differences their poisoning by anticancer and antibacterial drugs. Chem Biol 17: 421-433.
- Ishida R, Hamatake M, Wasserman RA, Nitiss JL, Wang, JC, et al. (1995) DNA topoisomerase II is the molecular target of bisdioxopiperidine derivatives ICRF-159 and ICRF-193 in Saccharomyces cerevisiae. Cancer Res 55: 2299-2303.
- Chen AY, Liu LF (1994) DNA topoisomerase essential enzymes and lethal targets. Annu Rev Pharmacol Toxicol 34: 191-218.
- Roy C, Brown DL, Little JE, Valentine BK, Walker PR, et al. (1992) The topoisomerase II inhibitor teniposide (VM-26) induces apoptosis in unstimulated mature murine lymphocytes. Exp Cell Res 200: 416-424.
- Endicott J, Ling V (1989) The biochemistry of P-glycoprotein-mediated multi drug resistance. Annu Rev Biochem 58: 137-171.
- Issacs R, Harris AL, Hickson ID (1996) Regulation of the human topoisomerase II a gene promoter in confluence-arrested cells. J BioI Chern 271: 16741-16747.
- Goswani PC, Roti Roti JL, Hunt CR (1996) The cell cycle-coupled expression of topoisomerase II during S phase is regulated by mRNA stability and is disrupted by heat shock or ionizing radiation. Mol Cell BioI 16: 1500-1508.
- Nakajima T, Ohi N, Arai T, Nozaki N, Kikuchi A, et al. (1995) Adenovirue E1A-induced apoptosis elicits a steep decrease in the topoisomerase II a level during the latent phase. Oncogene 10: 651-662.
- Nanbu T, Takahashi K, Murray JM, Hirata N, Ukimori S, et al. (2013) Fission Yeast RecQ Helicase Rqh1 Is Required for the Maintenance of Circular Chromosomes, Mol Cell Biol 336: 1175-1187.
- Nakano A, Masuda K, Hiromoto T, Takahashi K, Matsumoto Y, et al. (2014) Rad51 dependent aberrant chromosome structures at telomeres and rDNA activate the Spindle assembly checkpoint. Mol Cell Biol 34: 1389-1397.
- Pitt CW, Cooper JP (2010) Pot1 inactivation leads to rampant telomere resection and loss in one cell cycle. Nucleic Acids Res 38: 6968- 6975.
- Wang X, Baumann P (2008) Chromosome Fusions following telomere loss are mediated by single-strand annealing. Mol Cell 31: 463-473.
- Smith CL, Matsumoto T, Niwa O, Klco S, Fan JB, et al. (1987) An electrophoretic karyotype for Schizosaccharomyces pombe by pulsed field gel electrophoresis. Nucleic Acids Res 15: 4481-4491.
- Parsons AB, Brost RL, Ding H, Li Z, Zhang C, et al. (2004) Integration of Chemical-genetic and genetic interaction data link, bioactive compounds to cellular target pathways. Nat Biotechnol 22: 62-69.
- Yoshida S, Anraku Y (2000) Characterization of staurosporine-sensetive mutants of Saccharomyces Cerevisiae: Vacuolar functions affect staurosporine sensitivity. Mol Gen Genet 263: 877-888.
- Dawson K, Toone WM, Jones N, Wilkinson CR (2008) Loss of Regulators of Vacuolar ATPase Function and Ceramide Synthesis Results in Multidrug sensitivity in Schizosaccharomyces Pome. Eukaryot Cell 926-937.
- Baumann P, Cech TR (2000) Protection of telomeres by the Ku protein in ï¬ssion yeast. Mol Biol Cell 11: 3265–3275.
- Wang X, Baumann P (2008) Chromosome fusions following telomere loss are mediated by single-strand annealing. Mol Cell 31: 463-473.
- Sugawara N (1988) DNA sequences at the telomeres of the ï¬ssion yeast S. pombe. Ph.D. dissertation. Harvard University, Cambridge, MA.
- Nakamura TM, Cooper JP, Cech TR (1998) Two modes of survival of ï¬ssion yeast without telomerase. Science 282: 493-496.
- Tay Z1, Eng RJ, Sajiki K, Lim KK, Tang MY, et al. (2013) Cellular Robustness Conferred by Genetic Crosstalk Underlies Resistance against Chemotherapeutic Drug Doxorubicin in Fission Yeast. 8: e55041.