Research Article
Strain Improvement and Characterization of Lipase Produced by Pseudomonas Sp.
M.B. Sulochana*, Arunasri R, Parameshwar A.B and Mohan Reddy K
Corresponding author: M.B. Sulochana, Department of Post Graduate Studies and Research in Biotechnology, Gulbarga University, Gulbarga 585106, Karnataka, India,; E-mail: mbsulochana@rediffmail.com
Citation: Sulochana MB, Arunasri R, Parameshwar AB, Mohan Reddy K. Strain Improvement and Characterization of Lipase Produced by Pseudomonas Sp.. J Cell Sci Molecul Biol. 2014;1(1): 101.
Copyright © 2014 M.B. Sulochana et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Cell Science & Molecular Biology | Volume: 1, Issue: 1
Abstract
The improvement of microbial strains offers the greatest opportunity for the cost reduction without significant capital investment. The purpose of the present investigation is to enhance the production by subjecting the indigenous lipase producing strain Pseudomonas aeruginosa RAS-4 to strain improvement by using UV light as a physical mutagen and immobilization. Strain improvement studiesshowed that mutation was the best compared to immobilization technique showing 108% higher than the parent strain using UV light as a mutagenic agent. The molecular weight of lipase was determined to be 42 kDa. Localization of the lipase specific gene was carried out by plasmid curing and acridine orange proved to be the best curing agent revealing that the lipase gene is associated with the genomic DNA.
Keywords: Strain improvement; UV; immobilization
Introduction
Lipase enzyme plays a vital role in many food, dairy, leather, paper,pharmaceutical, detergent, textile and cosmetic industries. Lipases are secreted by microorganisms like bacteria, yeasts, molds and a few protozoa. The production of lipase by microorganisms depends largely on the species, strains and culture conditions. Microbial lipases are diverse in their enzymatic properties, substrate specificity and are usually more thermo stable than animal or plant lipases. Research on lipase production has intensified in recent years due to its potential application in various industrial processes. There are several reports on isolation of lipase producing microorganisms and the effect of nutritional factors on their growth and lipase production [1] determined the lipase activity by a rhodamine triglycerideagarose assay. Several species of Pseudomonas have been reported to produce this enzyme [2]. However, no reports are available on the production of lipase by mutated species of Pseudomonas. Therefore, the present study has been undertaken to study the efficacy of lipase production after inducing mutation in Pseudomonas sp. and also by immobilization technique. Quantitative enhancement could be obtained by strain improvement as the quantities produced by wild strains are low [3]. Improvement of the commercial applicability of microbial strains has been practiced for centuries [4]. Industrial strain improvement plays a central role in the commercial development of microbial fermentation processes. Meanwhile, strain improvement of the enzyme producer offers the greatest opportunity for cost effective production without significant capital outlay [5].
Materials and Methods
Collection of samples
Sterilized polythene bags were used for the aseptic collection of soil samples contaminated with oil and grease from different areas of oil mills of Gulbarga and Raichur regions. Samples were brought to laboratory and processed within 4 hours.
Isolation of the isolates
Pseudomonas Cetramide agar was used as a selective media for the isolation of Pseudomonas Identification of the colonies was done by the presence of bluish green pigmentation -pyocyanin [6].
Selective medium for the screening of lipase
Tributyrin agar containing 1.3% (w/v) nutrient broth was used as selective medium for the screening of lipase producing strains [7] and further screening was done for the rapid detection of lipase on Rhodamine B-Olive oil agar plates containing Rhodamine B .001% (W/V), agar 2%, nutrient broth 1.5% and olive oil 2.5% in distilled water (pH-7). Lipase production was monitored by irradiating plateswith conventional UV light at 350 nm [8].
Growth and Enzyme profile
Growth and enzyme profile of the enzyme was carried out using basal medium which consisted of Peptone - 10g, Beef extract - 3g, NaCl - 5g, Olive oil 1%, prepared in 1000 ml distilled water with pH – 8, temperature – 35°C and incubated for 36 h at 150 rpm. After incubation the samples were drawn and centrifuged at 1000 rpm for 10 min at 4°C. Cells were separated and culture filtrate was used for the assay of enzyme activity. The production of lipase and the growth was estimated at regular intervals of 4 h up to 48 h.
Enzyme assay
Lipase activity was measured according to [9], with little modification using p-nitrophenol palmitate (PNPP). One volume of 16.5mM PNPP (2-propanol) and 9 volumes of 50 mM Tris HCl buffer (pH-8) containing 0.4% Triton X-100 and 0.1% gum Arabic. The reaction was started by adding appropriate amount of enzyme solution of 100 μl and absorbance produced due to p-nitrophenol was determined at 410nm.
Phylogenetic analysis
The Pseudomonas sp. which showed maximum activity of lipase was characterized up to species level by molecular characterization. The strain was then subjected to 16S rRNA sequencing and was compared with the known sequences. The phylogenetic tree was constructed using the aligned sequences of similar species using Bioinformatics tool (www.ncbi.nim.nih/blast).
Strain improvement
The characterized strain of Pseudomonas sp. was subjected for strain improvement by mutational and immobilization studies. It was subjected to UV irradiation to induce mutations for the better yield of lipase according to the method of Saha and Bhattacharya [10]. The bacterial suspension of the parent strain was irradiated by using a 15 W Philips UV lamp for different time intervals ranging from 5 min to 30 min at a distance of 5cm to 20 cm away from the UV source. The irradiated suspension was protected from light by covering with black polythene bags in order to minimize any photo reactivation effects. This UV treated mutant isolates were then inoculated on tributyrin agar at 37°C for 24 h and the formation of zone around the colonies were confirmed as lipase production by the mutant strains.
Immobilization
Immobilization was carried out by entrapment of cells using Sodium alginate and calcium chloride [9]. Production of lipase by Pseudomonas sp. with the immobilized cells was carried out by using the immobilized beads into a 50 ml of production medium in 250 ml of Erlenmeyer’s flask, for 24-48 hours in batch fermentation for the lipase production. For the evaluation of bead stability, immobilization was carried out with different concentrations of sodium alginate from 2- 5% and CaCl2 solution from 0.1 M – 0.4 M. Then the beads were inoculated into the production medium for lipase production and the enzyme activity of free cells and immobilized cells were compared.
Determination of molecular weight of lipase
Molecular weight of lipase was determined by using SDS-PAGE, according to the method of Laemmli (1970). 100 μg of lipase was dissolved in 10 μl of sample buffer 1% SDS, 5% Mercaptoethanol, 0.003% Î’romophenol blue and 10% Glycerol in 0.063 M Tris HCl pH 6.8 were boiled at 100°C for 3 min. 12.5% separating gel was poured and allowed it to solidify. 5% stacking gel was poured on separating gel and comb was inserted and allowed to solidify and the sample was electrophoresed with 0.025 M Tris HCl electrode buffer containing glycine and 0.1% SDS. After the electrophoresis the gel was stained with Coomaise Brilliant Blue R-250 and left overnight to visualize the protein bands.
Zymogram
The native polyacrylamide gel electrophoresis (PAGE) of lipase was carried out as described by [11]. The electrophoresed gel was placed in petriplate containing 2% agar, 3% olive oil and 0.001% Rhodamine B and was incubated at 37°C for 3-4 h. The lipase activity was detected by observing zone of hydrolysis corresponding to the position of band showing lipase activity.
Determination of lethal dose of curing agents for Pseudomonas
Plasmid curing experiment was carried out in order to localize the lipase specific trait in the genome of the Pseudomonas sp. using ethidium bromide and acridine orange as curing agents. The lethal dose for Pseudomonas sp. was determined, in the nutrient broth containing acridine orange and simultaneously the experiment was run on nutrient agar plates impregnated with acridine orange. 0.1 ml of 20 h old Pseudomonas sp. culture was inoculated into 50 ml of sterilized nutrient broth (pH-7) supplemented with different concentrations of curing agents ranging from 1- 10 μg/ml were incubated at 37°C for 24 h and kept on an orbital rotatory shaker at a speed of 150 rpm. The concentration of curing agent at which the Pseudomonas sp. grew with 0.25-0.3 at OD- 660 after 24 h of incubation, this was selected as the concentration for the elimination of plasmid from Pseudomonas sp.
Results and Discussion
Isolation and characterization of the strain was carried out by following [4], Pseudomonas sp. is a gram negative, non-spore forming, non-motile. The microscopic and biochemical tests are as shown in Table 1.
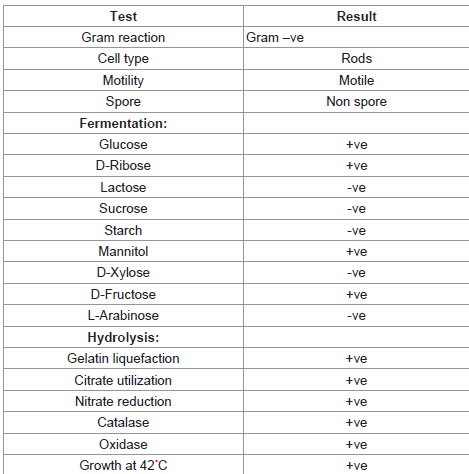
Table 1: Morphological and Biochemical characteristics of Pseudomonas sp.
Screening of lipase
The qualitative assay was done for screening of lipase on Tributyrinagar plates and was confirmed by the presence of clear zone of around the colonies as shown in Figure 1. Similar observation was reported by [12]. True lipase producers were screened on Rhodamine B olive oil; the presence of bright orange fluorescence halo around the colony indicated the lipolytic activity.
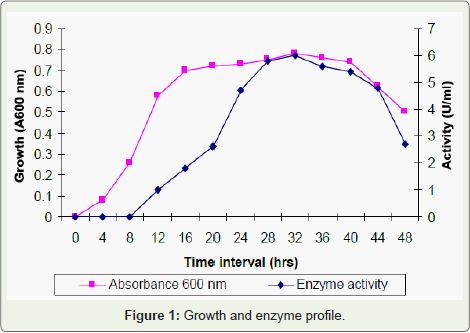
Figure 1: Growth and enzyme profile.
Growth and lipase production profile
Growth of the strain began after 4h and reached the maximum up to 48h and then gradually decreased. Simultaneously the production of enzyme started after 8h and reached the maximum at 32h with the enzyme activity of 6 U/ml and there after decreased as shown in Figure 2. These observations corroborate the findings on B. subtilis [13], P. aeruginosa [1] where maximum lipase activity was produced in the late exponential growth phase and then decreased as the growth decreased. The decline in the extracellular lipase may be due to the increase in oleic acid concentration.
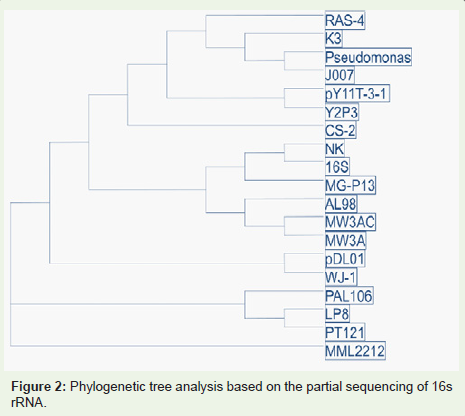
Figure 2: Phylogenetic tree analysis based on the partial sequencing of 16s rRNA.
Phylogenetic analysis
The 16S rRNA sequence of Pseudomonas sp. was compared with NCBI database and the phylogenetic tree was constructed which is as shown in Figure 3. This strain showed homology of 99.9% with P. aeruginosa K3 and was identified and designated as P. aeruginosa RAS-4.
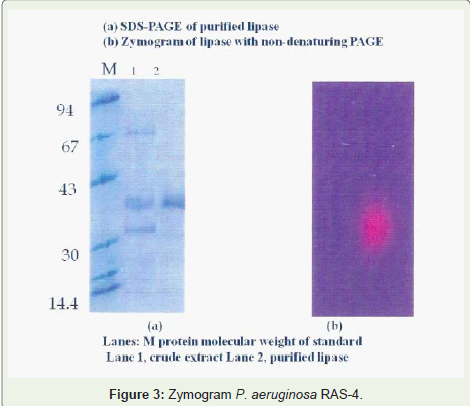
Figure 3: Zymogram P. aeruginosa RAS-4.
Strain Improvement
A potent mutated bacterial strain was isolated and characterized from oil mills and was designated as P. aeruginosa RAS-4, which produced maximum enzyme activity of 12 U/ml. The lipolytic activity was screened by rapid plate method and the appearance of clear zone of hydrolysis on tributyrin and orange fluorescence on Rhodamine agar was observed around the colonies. [3] and [5] reported that the improvement of the microbial strain offers the greatest opportunity for the cost reduction without significant capital investment. Survival data of Pseudomonas sp. on UV treatment are similar to the results of [51] as shown in Table 2.
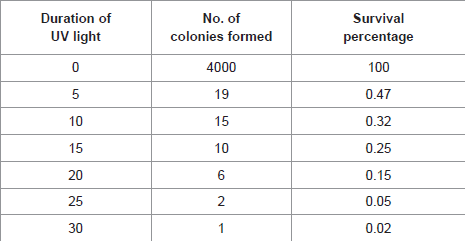
Table 2: Morphological and Biochemical characteristics of Pseudomonas sp.
Immobilization - Effect of sodium alginate concentration
To determine the optimum concentration of sodium alginate as an immobilization matrix, sodium alginate of 2% to 5% were prepared and bacterial suspension was added to it. The beads thus formed were inoculated into the production medium. For the evaluation of bead stability, the immobilization procedure was carried out with different concentrations of CaCl2 solution from 0.1 M – 0.4 M. The beads werethen inoculated into the production medium for lipase production and the enzyme activity of free cells and immobilized cells were compared as shown in Table 3.
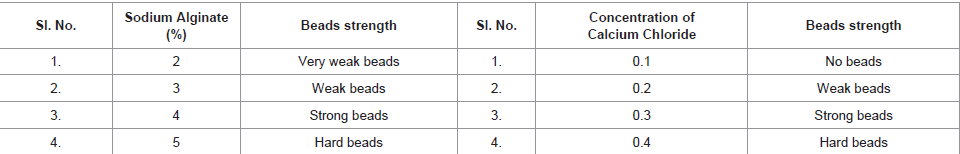
Table 3: Optimization of sodium alginate and Calcium Chloride.
Determination of Molecular Weight and Zymogram
The molecular weight of the enzyme was determined by SDSPAGE along with standard markers and the molecular weight of lipase was found to be 42 kDa as shown in Figure 4. The molecular weight of the homogenous lipase was determined about 42 kDa by SDS-PAGE for Mucor sp. [17]. The lipase produced by Pseudomonas aeruginosa YS -7 showed 40 kDa as reported by [18], a single band was observed in native gel under 350nm UV light that confirmed the homogencity of the lipase and showed olive hydrolysis properties during running the zymogram [19] detected 50 kDa purified lipase from P. flurosens by activity staining on olive oil and Rhodamine B similar finding were also reported by [20] from Mucor meihei, Pseudomonas, Burkholderia multivorans and P. aeruginosa on the gel by zymography.
Localization of Lipase Gene by Plasmid Curing
The localization of the lipase specific trait in Pseudomonas aeruginosa RAS-4 genome was made by indirect method of plasmid elimination experiment. 7ug/ml acridine orange was the best concentration and plasmid was eliminated at 8th generation. Based on these results acridine orange was found to be the best curingagent. The lipase activity was not altered when the plasmid of isolate RAS-4 was completely eliminated, based on all the above results it is confirmed that the lipase gene is located in chromosomal DNA of the isolate RAS-4 but not in plasmid. [21] Used acridine orange, ethidium bromide and SDS as a curing agent on phenotypic stability in Pseudomonas putida degrading E-carpolactum. Curing analysis of P. aeruginosa was carried out by ethidium bromide, acridine orange, novobiocin and SDS as per the reports of Edward Raja and Selvam [22].
Summary
A rapid plate method was done for screening of lipase on Tributyrin agar plates and was confirmed by the presence of clear zone of hydrolysis of 4.2cm on tributyrin and orange fluorescence on Rhodamine agar. 16S rRNA showed 99.9% homology with Pseudomonas aeruginosa K3 strain and consequently the isolate was identified and designated as Pseudomonas aeruginosa RAS-4. Strain improvement studies showed, mutation was the best when compared to the immobilization technique showing 108% higher yield than the parent strain using UV light as a mutagenic agent. The molecular weight of lipase from non-mutated was determined to be 42 kDa. Localization of lipase specific gene was carried out by plasmid curing and acridine orange proved to be the best curing agent revealing that the lipase gene is associated with the genomic DNA. Isolation of Pseudomonas aeruginosa RAS-4 a potential strain producing lipase is an important contribution, however, the mutant strain RAS-4 isolated from parent is comparatively efficient producer of lipase.
References
- Kamini NR, Fuji T, Kurosa T, Ifeuji H (2000) Production, purification and characterization of an Extracellular lipase from yeast Cryptococcus sp. S-2. Process Biochemistry 36: 317-324.
- Suzuki T, Mushiga Y, Yamane T, Shimizu S (1998). Mass production of lipase by fed batch culture Psedomonas fluorescence. Applied Microbiology and Biotechnology 27 : 417-422.
- Bapiraju KVSN, Sujatha P, Ellaiah P, Ramanna T (2004) Mutation induced enhanced Biosynthesis of lipase. African J Biotechnolo 3: 618-621.
- Parekh S, Vinci VA, Strobel RJ (2000) Improvement of microbial strains and fermentation processes. Appl Microbiol Biotechnol 54: 287-301.
- Stanbury PF, Whitaker, A, Hall SJ (1995). Media for industrial fermentations. In: Principles of fermentation technology. (ed. P. F. stanbury, A Whitaker and S. J. Hall) Elsevier Science Ltd., Pergamon, Oxford, New York, Tokyo. 93-122.
- Bergey’s Manual of systematic Bacteriology (1984). William and Wilkins Publications 1.
- Lawrence RC, Frayer TF, Reiter B (1967) Rapid method for the quantitative estimation of microbial lipase. Nature 213: 1264-1265.
- Kouker A, Jaeger KE (1987) specific and sensitivity plate assay for bacterial lipases. Appl Environ Microbil 53: 211-213.
- Kordel M, Hofman B, Schomburg D, Schimd RD (1991) Extracellular lipase of Pseudomonas sp. Strain ATCC 21808 Purification and characterization and preliminary X-ray diffraction data. J Bacteriol 173: 4836-4841.
- Shah and Bhattacharya (1990) Strain improvement mutagenesis and random screening procedure for Rhizopus oryzae IITKG-1 proceeding of international symposium on industrial Biotechnology, Department of Microbiology. Osmania University, Hyderabad.
- Johanson A, Flink M (1986) Influence of alginate properties and gel reinforcement on fermentation characteristic of immobilized yeast cells. Enzyme Microb Technol 8: 737- 742.
- Andrew’s AT (1986) Electrophoresis theory, techniques and biochemical and clinical applications, 2nd edition Clarendon Press, Oxford.
- Senthilkumar R, Selvakumar G (2008) Isolation and characterization of extracellular lipase producing Bacillu. sp. SS-1 from slaughter house soil. Advanced Biotechnol PP 24- 25.
- Lesuisse E, Schnak K, Colson (1993) Purification and preliminary characterization of extracellular lipase of Bacillus subtilis 168, an extremely basic pH tolerant Enzyme. Eur J Biochem 216: 155-160.
- Gilbert EJ, Drozo JW, Jones CW (1991) Physiological regulation and optimization of lipase activity in Pseudomonas aeruginosa. J Gen Microbiol 137: 2215-2221.
- Jayati Ray Dutta, Rintu Banarjee (2006) Isolation and Characterization of newly isolated pseudomonas mutant for protease production. An International Journal 49: 37-47.
- Abbas H, Hiol A, Deytris V, Comeau L (2002) Isolation and characterization of extracellular lipas from Mucor sp.strain isolated from palm fruit.
- ShabtaiY, Mishne ND (1992) production, purification and properties of lipase from a bacterium(Pseudomonas aeruginosaYS-7) capable of growing in water restricted environments. App Environ Microbial 58: 174-180.
- Lee Y, Rhee JS (1993) Production and partial purification of lipase from Pseudomonas putida 3SK. Enzyme MicrobTechnol.
- Singh R, GuptaN, Goswami VK, Gupta R (2006) A simple activity staining protocol for lipase and esterases. Appl Microbiol Biotechnol 70: 679-682.
- Kulkarni RS, kanekar PP (1997) Effects of some curing agents on phenotypic stability in Pseudomonas putida degrading e-caprolactum. World Journal of Microbiology and Biotechnology. 14: 255-257.
- Edward Raja C, Selvam GS (2009) Plasmi profile and curing analysis of Pseudomonas aeruginosa as a metal resistant. Intl J Environ Sci Tech 6:259-266.