Review Article
Understanding the Mechanism of Breast and Prostate Cancer Cell Killing by Diallyldisulfide – An Over View
Sujatha P1, Siddhartha V Talluri2, Suma M Natraj1, Gowthamarajan Kuppusamy3#, Subbarao V Madhunapantula4#*
Corresponding author: Dr. SubbaRao V. Madhunapantula, Associate Professor, Center of Excellence in Molecular Biology and Regenerative Medicine, Department of Biochemistry, JSS Medical College, JSS University, Mysuru, Karnataka, India, Pin code: 570 015, Mobile: +91-810-527-8621; E-mail: madhunapantulas@yahoo.com
Citation: Sujatha P, Siddhartha VT, Suma MN, Gowthamarajan K, Subba Rao VM. Understanding the Mechanism of Breast and Prostate Cancer Cell Killing By Diallyldisulfide – An Over View. J Chem Applied Biochem. 2015;2(1): 112.
Copyright © 2015 SubbaRao V. Madhunapantula et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Submission: 20/08/2015; Accepted: 28/09/2015; Published: 03/10/2015
Abstract
Carcinomas of Breast and Prostate are the second leading causes of deaths due to cancers, respectively, in women and men worldwide. Incidence and mortality rates due to these cancers have been increasing at an alarming rate each year. Lack of effective early diagnostic tools and potent therapeutic agents are the key contributing factors for global burden of these cancers. Therefore, effective, less toxic and affordable treatment agents are urgently required, and the sulfur containing compounds from garlic represent one such group of agents with better therapeutic efficacy and acceptable safety profiles. Among various sulfur containing compounds isolated from garlic (Allium sativum L) and tested for anti-cancer properties in vitro and in preclinical animal models, the diallyl disulfide (DADS) and diallyl trisulfide (DATS) have been reported to exhibit better therapeutic properties. Even though prior studies have shown marginally better therapeutic efficacy of DATS compared to DADS, the latter was preferred due to its low cost, more abundance and easy availability. Anti-cancer properties of DADS have been tested in vitro using cell lines representing carcinomas of prostate and breast. Results of these studies proposed that DADS induced cell death is due to the (a) upregulation of apoptosis proteins; (b) downregulation of cell proliferation molecules, (c) arrest of cell cycle progression through the induction of key proteins participating in the DNA synthesis and replication and (d) inhibition of cell migration and metastasis. However, it is not fully understood how DADS inhibit tumor cell metastasis. Likewise it is also not fully known how DADS could retard the growth of a range of tumors? Is there any common mechanism operating in these tumors, which is regulated by DADS? Therefore, in this review article we determined to address these gaps by a thorough search of published findings
Keywords: Garlic; DADS; DATS; Breast cancer; Prostate cancer; Anti-cancer activity
Introduction
Breast and prostate cancers are the major culprits affecting the health of women and men globally. According to IARC-GLOBOCAN, while carcinomas of breast accounts to 1.67 million new cases and 0.521 million deaths, the carcinomas of prostate represent 1.1 million new cases and 0.307 million deaths worldwide in the year 2012 [1]. Recent trends assessing the incidence and mortality rates shows that these numbers rapidly increase in the near future if preventive, and curative measurements are not immediately implemented. Highlighting these points, recently, International Agency for Research on Cancer (IARC) announced that there is “an urgent need in cancer control today to develop effective and affordable approaches to the early detection, diagnosis, and treatment, especially for individuals living in developing countries” [1]. However, to date, no effective cancer preventive or treatment agents are available to reduce the incidence and treatment burden of breast and prostate cancers. Moreover, resistance to several of existing therapeutic agents has been reported in breast and prostate cancers, necessitating the requirement of multi-target inhibitors with minimal toxicity [2,3]. But, no such multi-target agent with good potency and better safety profile exists. In addition, since majority of chemotherapeutic agents are synthetic and/or semisynthetic compounds with poor pharmacokinetic and safety profile, identifying a therapeutic agent form natural sources is important.
Garlic (Allium sativum) and garlic derived dietary products have gained importance due to various health beneficial effects such as (a) Reduction of cholesterol thereby cardiovascular diseases; (b) Prevention and treatment of cancer; (c) Boosting of the immune system; (d) Detoxification of toxic compounds thereby restoring physical strength; (e) Protection from radiation damage [4-9]. Strengthening these observations, several clinical trials conducted using garlic and garlic extracts have also shown hypocholesterolemic and cancer preventive effects [8,10,11]. According to a recent report China and India are the major producers of garlic and garlic derived foods worldwide with China producing >70% of total garlic produced in the world. Various garlic based food products are categorized in to (a) Essential oils; (b) Macerates; (c) Powder; and (d) Extract [12]. Essential oils are produced by steam distillation of garlic and usually contain a variety of sulfides that include DADS and DATS but devoid of allicin and water-soluble compounds [12]. Garlic oil macerates are made of whole garlic cloves ground in to vegetable oil (sesame oil / sunflower oil). Even though part of the alliin is converted to allicin, the amount of allicin in the final product is usually very minimal due to its labile nature. Therefore, the finished product contains allicin-decomposed compounds such as dithiins, ajoene and sulfides etc. Garlic powder is produced by drying the sliced or crushed garlic followed by pulverization in to powder [12,13]. Although it is believed to contain all the major constituents of intact garlic, the proportion and amount of each ingredient differ from the original due to processing. For example, the dehydrated garlic powder should contain about 2.5 mg/g alliin, however, the garlic powder usually contain < 1 mg/g of alliin indicating that majority of alliin is lost during preparation. Another product of garlic is aged garlic extract (AGE) [14]. AGE is prepared by soaking the whole or sliced garlic cloves in water or dilute alcohol for several months followed by separation of the solution and concentration to obtain a thick extract [14]. The extract contains very high amounts of water-soluble sulfur containing compounds that include S-allyl cysteine (SAC) and S-allyl mercaptocysteine (SAMC) [14]. Efficacy of these garlic products varies from the method of preparation and storage. For example, a set of clinical studies reported between the years 1990 and 2000 have demonstrated that not all garlic preparations are beneficial, but only a set of preparations provide health promoting effects, and the variability in efficacy results in the published studies could be due to the differences in the method of isolation, fractionation and quality of preparations [15]. Therefore, it is important to understand the chemistry of garlic constituents, and their effect on various cellular events such as cell proliferation, apoptosis and metastasis.
Synthesis and Anticancer Properties of DADS
Biosynthesis of DADS: Diallyldisulfide, chemically known as 4,5-dithia-1,7-octadiene (DADS), is one of the major phytoconstituents isolated from garlic [15]. Garlic is a widely used herbal supplement and spice with sulfur containing compounds γ-glutamylcysteines, and cysteine sulfoxides [15]. DADS is a oil soluble compound produced from the decomposition of Allicin. Allicin is produced from the Allycysteine sulfoxide (ACS), also known as Alliin. Alliin is the most abundant sulfur-containing compound that represents up to 80% cysteine sulfoxides [16]. When garlic is crushed, alliinase enzyme gets released and promotes the synthesis of Alliin, which further gets converted in to sulfenic acid. The sulfenic acid thus produced spontaneously reacts with each other to form unstable thiosulfinate, also known as Allicin [16]. Allicin further breaks down in vitro to form diallyl sulfide (DAS), diallyl disulfide (DADS) and diallyl trisulfide (DATS) (Figure 1) [17].
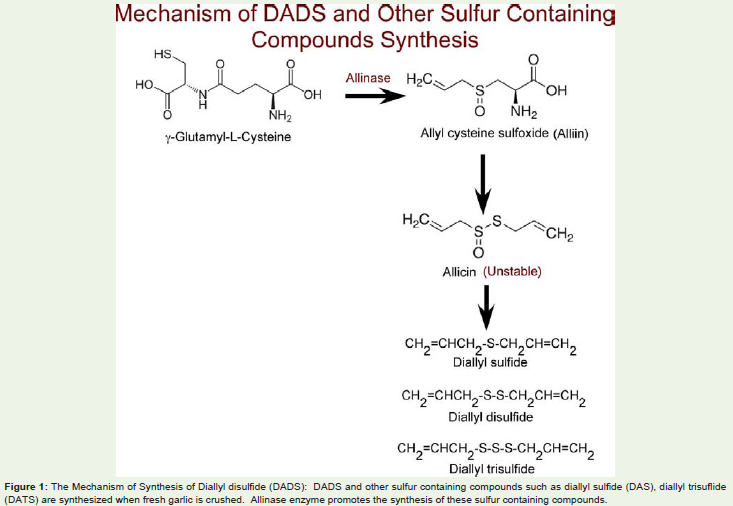
Figure 1: The Mechanism of Synthesis of Diallyl disulfide (DADS): DADS and other sulfur containing compounds such as diallyl sulfide (DAS), diallyl trisuflide (DATS) are synthesized when fresh garlic is crushed. Allinase enzyme promotes the synthesis of these sulfur containing compounds.
DADS as an anti cancer agent: Several in vitro culture models as well as preclinical experimental animal studies have shown that DADS could inhibit the proliferation of cancer cells thereby promote tumor regression [18,19]. Mechanistically, DADS has been reported to (a) activate enzymes that detoxify carcinogens [20]; (b) suppress the formation of DNA adducts [21,22]; (c) exhibit antioxidant effects [23,24]; (d) induce cell-cycle arrest and trigger apoptosis [25,26]; (e) promote transformation of cancer cells to normal cells [27]; (f) trigger histone modification in such a way that augment the tumor suppressor expression [28-30]; and (g) inhibit the process of angiogenesis, invasion and metastasis [31]. In this article we review the recent reports showing the anticancer properties of DADS for inhibiting the growth of breast and prostate cancers.
DADS for inhibiting breast cancers: Growth inhibitory properties of DADS and DATS against human breast cancer cell lines T47D and MCF-7, which express estrogen and progesterone receptors (ER and PR) but lacking human epidermal growth factor receptor (HER2), were reported by Lea M et al., in 1997 and Hahm ER and Singh SV in 2014, respectively [32-34]. Authors have reported that thymidine incorporation in to the DNA of these cell lines was inhibited by 1 to 2 mM DADS demonstrating the anti-proliferative effects of this organosulfur compound [32]. Mechanistically, treatment of these cells with DADS or its metabolite allylmercaptan inhibited histone deacetylase (HDAC) activity thereby enhanced the levels of acetylated histones H4 and H3 [29]. Supporting this initial observation, a recent article provided further evidences by reporting HDAC inhibitory activity leading to elevated acetylated H4 levels when MCF-7 cells were exposed to DADS [29]. Targeted inhibition of HDAC by DADS induced apoptosis by increasing the expression of apoptotic proteins such as caspase-3 and Bax while decreasing anti-apoptotic protein Bcl2 [29,35]. A separate study has shown that treatment of MCF-7 cells with 200μM DADS for 6h increased apoptosis and inhibited proliferation by decreasing phosphorylated extracellular signal regulated kinase (pERK) while promoting the levels of active phosphorylated c-Jun N-terminal kinase (pJNK) and phosphorylated p38 (p-p38) proteins [34]. Mechanistically, exposure of breast cancer cells to DADS induced the expression of miR-34a thereby decreased SRC/Ras/ERK signaling [36]. Prior studies have shown that miR-34a controls cell proliferation and survival signaling cascades in cancers [37,38]. In addition to the inhibition of proliferation signaling pathways, DADS and the trisulfide version DATS have also been reported to increase reactive oxygen species (ROS) to trigger apoptosis in MCF-7 breast cancer cells [39]. Interestingly, a recent study for the first time reported that DADS and DATS could inhibit ERα as well as peptidylprolyl cis-trans isomerase (Pin-1) expression in MCF-7 and T47D cell lines to promote cell death [33]. Furthermore, in addition to controlling the proliferation and apoptotic pathways, DADS has been reported to halt the TNF-α induced release of CCL2. CCL2, also known as monocyte chemotactic protein (MCP-1), release by tumor cells enables homing of mononuclear cells in the vicinity of tumor tissue, which subsequently get transformed in to pro-oncogenic tumor-associated macrophages (TAMs) [40,41]. Therefore, agents like DADS, that inhibit the release of CCL2 from tumor cells promote tumor cell degradation. In conclusion, the anti-breast cancer effects of DADS are mediated by inhibiting the proliferation by preventing the signaling cascades while inducing the pro-oncogenic apoptotic pathways and transformation of mononuclear cells in to TAMs (Figure 2).
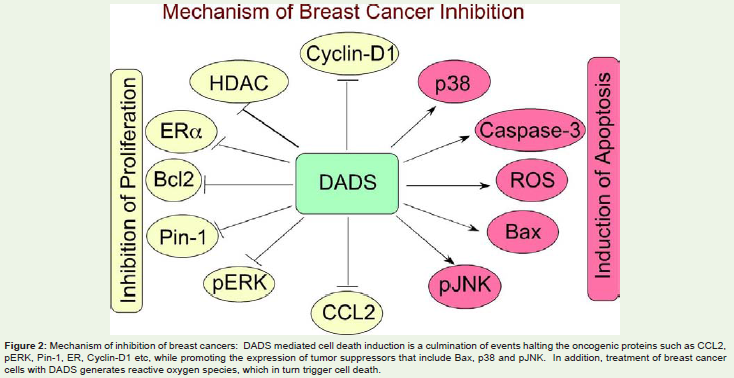
Figure 2: Mechanism of inhibition of breast cancers: DADS mediated cell death induction is a culmination of events halting the oncogenic proteins such as CCL2, pERK, Pin-1, ER, Cyclin-D1 etc, while promoting the expression of tumor suppressors that include Bax, p38 and pJNK. In addition, treatment of breast cancer cells with DADS generates reactive oxygen species, which in turn trigger cell death.
DADS for inhibiting prostate cancers: Arunkumar A et al., in 2005 have demonstrated anti-prostate cancer effects of DADS by comparing the levels of incorporation of [3H]thymidine and DNA fragmentation in PC-3 cell line [42]. Authors have reported that 50 and 100 μM DADS could inhibit cell growth by decreasing the [3H] thymidine incorporation and by inducing apoptosis as evidenced by elevated number of ethidium bromide stained cells, and increased DNA breaks observed when analyzed by agarose gel electrophoresis [42]. Mechanistically, DADS and other sulfur containing compounds from garlic have shown to induce (a) cell cycle arrest in G2-M phase by inhibiting CDK1 [43]; (b) phosphorylation of Bcl2 thereby by reducing the levels of Bcl2:Bax interactions [44]; (c) promoting cleavage of procaspase-9 and -3 to trigger apoptosis [35] (Figure 3). Since members of Bcl family proteins regulate cell survival and apoptosis via modulating the levels of Bcl2, Bid and Bax, the effect of DADS and other sulfur containing compounds from garlic to decrease anti-apoptotic Bcl2 while increasing the pro-apoptotic Bax are significant [45]. Phosphorylation of Bcl2 is a key event in the regulation of apoptosis and is mediated by the activated JNK1 and 2 and ERK1/2 [45]. Since DADS and DATS have been shown to activate JNK1 and JNK2 as well as ERK1/2 proteins, it was postulated that these sulfur containing garlic compounds trigger the apoptosis by activating these proteins followed by inhibiting the pro-survival Bcl2 [45]. Supporting these observations, a separate study recently demonstrated that DADS could inhibit the growth of DU145 cells by modulating the levels of Bcl-2 and inhibitor of apoptosis protein (IAP) by promoting the phosophorylation of ERK, and c-Jun N-terminal kinase (JNK) [26]. Furthermore, the authors have shown that DADS could also stimulate the proteolytic activation of caspases, induce the expression of death-receptor 4 (DR4) and Fas ligand (FasL) proteins while decreasing the levels of Bid proteins [26]. Proof-of-principle studies using a specific JNK inhibitor, SP600125, significantly blocked DADS-induced-apoptosis [26]. However, the inhibitors of MAPK pathway such as U0126 had only marginal effects on Bcl2 phosphorylation, indicating that MAPK signaling has a minor role in controlling the Bcl2 expression and cellular apoptosis compared to JNK [26]. In addition to controlling the JNK and MAPK signaling, DADS could also control the induction of apoptosis by inactivating phosphatidylinositol 3-kinase (PI3K)/Akt pathway [26]. Since PI3KAkt signaling has been reported to control cancer cell survival by down-regulating apoptosis, inhibiting this signaling cascade by DADS could induce apoptosis and make the cells sensitive to various chemotherapeutic agents [46]. Furthermore, the docking studies using Ligand Fit model have confirmed that DADS could inhibit PI3K-Akt signaling by modulating the upstream IGF pathway [47].
Figure 3: Mechanism of inhibition of prostate cancers: DADS has been shown to trigger the inhibition of prostate cancers by down-regulating the expression/activity of Akt, CDK1, Bid, MMP-2 and MMP-9. The apoptosis induction due to DADS treatment is mediated by apoptotic proteins such as DR4, Fas ligand and promoting the release of calcium from endoplasmic reticulum.
Several other mechanisms have also been attributed to the DADS induced apoptosis in prostate cancers. For example, in one study DADS has been shown to promote histone acetylation thereby inhibit cell growth via induction of apoptosis [48]. However, it is not fully clear whether DADS induced histone acetylation is the result of inhibition of deacetylation of histones by halting the activity of HDACs or promoting the acetylation of histones via the activation HATs. More over, it is also not fully known how increased histone acetylation is promoting tumor cell apoptosis? Therefore, further studies are warranted to elucidate these mechanisms. Another study showed that DADS could raise the intracellular Ca2+ by promoting the Ca(2+) influx via phospholipase A(2)-sensitive channels as well as by activating calcium channels in endoplasmic reticulum [49]. Elevated intracellular calcium due to DADS treatment promoted cellular death in prostate cancers. Pretreatment with calcium chelater 1,2-bis(2-aminophenoxy)ethane-N,N,N’,N’-tetraacetic acid (BAPTA) partly reduced the cytotoxic effects of DADS indicating the role of elevated calcium levels in the induction of apoptosis in prostate cancers [49].
Tumor cell metastasis is another key event in the progression of cancers [50]. The process of metastasis involves (a) tumor cell dissociation from primary tumors; (b) invasion of blood vessel epithelium; (c) circulation in the blood (d) extravasation; and (e) formation of secondary tumors. Majority of cancer related deaths are due to aggressive metastatic stages. Therefore, therapeutically targeting these metastatic stages is highly significant [51]. Preliminary studies using DADS have shown the inhibition of invasiveness of prostate cancers cells [52]. Mechanistically, DADS repressed the expression of tight junction protein claudin, which has a key role in control and selectivity of paracellular transport [52]. In addition, DADS could also inhibit matrix metalloproteinase (MMP)-2 and -9 in LNCaP cells [52]. However, it is not fully known whether DADS could decrease tumor cell metastasis in other cancers.
Pharmacokinetic properties of Dads
Preclinical animal studies testing the efficacy of DADS for inhibiting the growth of xenografted breast- and colorectal cancers have shown a significant reduction in the tumor volume when DADS was given intraperitoneally [36,53]. For example, a recent study showed an about 60% tumor growth inhibition when 100mg/kg DADS was injected intraperitoneally in to mice bearing xenografted breast tumors [36]. Similarly, another study reported shrinkage of subcutaneous Colo-205 tumors when 6mg/kg DADS was injected intraperitoneally in to mice [53]. Even though these in vivo studies have reported the tumor inhibitory properties of DADS not much is known about the pharmacokinetic and pharmacodynamic studies of DADS. Recent studies determining the pharmacokinetic studies of DADS demonstrated the possible transformation of orally administrated DADS when injected in to rats [54]. For instance, a single oral administration of 200 mg/ kg DADS to rats showed a quick absorption and transformation into allyl mercaptan, allyl methyl sulphide, allyl methyl sulphoxide (AMSO) and allyl methyl sulphone (AMSO(2) [54]. These metabolites have been shown to reach peak maximum between 48-72h after the administration of DADS. Stability assessment studies have demonstrated that among these DADS derived metabolites, AMSO(2) was the most abundant and persistent metabolite and gets excreted in the urine [54]. Further experiments measuring the time kinetics of all these sulfur compounds using GCMS showed a rapid decline within the first week after administration and disappearance during the second week. Even though these potential metabolites are thought to be active in the target tissues, further studies are warranted to conclusively establish their effect on cancer cells.
Summary
In summary, DADS mediated cell growth inhibition is mediated by down-regulation of key oncogenic proteins expression and or activity while inducing the levels of tumor suppressors.
Future Directions
Even though the anti-cancer properties of DADS have been extensively studied, many unaddressed aspects still require further investigations. For example, it is not fully known whether DADS physically binds to any key cell proliferation regulator to inhibit its activity. In this regard studies should focus on co-crystallization methods to isolate protein-DADS complexes and check how this binding modulate the activity of the protein. Likewise, it is also not clear whether DADS has the ability to inhibit tumor cell metastasis when administered to animals suffering from metastatic tumor load. If inhibiting, what is the mechanism of metastasis suppression? Further studies are also required whether DADS synergizes with other sulfur containing compounds such as phenethylthiosulfate (PEITC) or any known anti-cancer agents? Experiments need to be performed in this direction to assess and identify appropriate synergistic agent and to check whether this combination potentially reduce the otherwise required high dose of DADS.
Acknowledgements
Authors would like to acknowledge JSS University for providing research support.
References
- Torre LA, Bray F, Siegel RL, Ferlay J, Lortet-Tieulent J, et al. (2015) Global cancer statistics, 2012. CA Cancer J Clin 65: 87-108.
- Yardley DA (2013) Drug resistance and the role of combination chemotherapy in improving patient outcomes. Int J Breast Cancer 2013: 137414.
- Semenas J, Allegrucci C, Boorjian SA, Mongan NP, Persson JL (2012) Overcoming drug resistance and treating advanced prostate cancer. Curr Drug Targets 13: 1308-1323.
- Nicastro HL, Ross SA, Milner JA (2015) Garlic and onions: their cancer prevention properties. Cancer Prev Res (Phila) 8: 181-189.
- Sultan MT, Butt MS, Qayyum MM, Suleria HA (2014) Immunity: plants as effective mediators. Crit Rev Food Sci Nutr 54: 1298-1308.
- Zhang CL, Zeng T, Zhao XL, Xie KQ (2013) Garlic oil attenuated nitrosodiethylamine-induced hepatocarcinogenesis by modulating the metabolic activation and detoxification enzymes. Int J Biol Sci 9: 237-245.
- Butt MS, Sultan MT, Iqbal J (2009) Garlic: nature's protection against physiological threats. Crit Rev Food Sci Nutr 49: 538-551.
- Butt MS, Sultan MT (2009) Green tea: nature's defense against malignancies. Crit Rev Food Sci Nutr 49: 463-473.
- Khanum F, Anilakumar KR, Viswanathan KR (2004) Anticarcinogenic properties of garlic: a review. Crit Rev Food Sci Nutr 44: 479-488.
- Bayan L, Koulivand PH, Gorji A (2014) Garlic: a review of potential therapeutic effects. Avicenna J Phytomed 4: 1-14.
- Rana SV, Pal R, Vaiphei K, Sharma SK, Ola RP (2011) Garlic in health and disease. Nutr Res Rev 24: 60-71.
- Amagase H, Petesch BL, Matsuura H, Kasuga S, Itakura Y (2001) Intake of garlic and its bioactive components. J Nutr 131: 955S-62S.
- Lawson LD, Hughes BG (1992) Characterization of the formation of allicin and other thiosulfinates from garlic. Planta Med 58: 345-350.
- Lawson LD, Gardner CD (2005) Composition, stability, and bioavailability of garlic products used in a clinical trial. J Agric Food Chem 53: 6254-6261.
- Omar SH, Al-Wabel NA (2010) Organosulfur compounds and possible mechanism of garlic in cancer. Saudi Pharm J 18: 51-58.
- Borlinghaus J, Albrecht F, Gruhlke MC, Nwachukwu ID, Slusarenko AJ (2014) Allicin: chemistry and biological properties. Molecules 19: 12591-12618.
- Block E (1985) The chemistry of garlic and onions. Sci Am 252: 114-119.
- Yi L, Su Q (2013) Molecular mechanisms for the anti-cancer effects of diallyl disulfide. Food Chem Toxicol 57: 362-370.
- Yun HM, Ban JO, Park KR, Lee CK, Jeong HS, et al. (2014) Potential therapeutic effects of functionally active compounds isolated from garlic. Pharmacol Ther 142: 183-195.
- Nkrumah-Elie YM, Reuben JS, Hudson AM, Taka E, Badisa R, et al. (2012) The attenuation of early benzo(a)pyrene-induced carcinogenic insults by diallyl disulfide (DADS) in MCF-10A cells. Nutr Cancer 64: 1112-1121.
- Sapkota M, Hottor TK, DeVasure JM, Wyatt TA, McCaskill ML (2014) Protective role of CYP2E1 inhibitor diallyl disulfide (DADS) on alcohol-induced malondialdehyde-deoxyguanosine (M1dG) adduct formation. Alcohol Clin Exp Res 38: 1550-1558.
- Lin JG, Chen GW, Su CC, Hung CF, Yang CC, et al. (2002) Effects of garlic components diallyl sulfide and diallyl disulfide on arylamine N-acetyltransferase activity and 2-aminofluorene-DNA adducts in human promyelocytic leukemia cells. Am J Chin Med 30: 315-325.
- Kim JM, Chang HJ, Kim WK, Chang N, et al. (2006) Structure-activity relationship of neuroprotective and reactive oxygen species scavenging activities for allium organosulfur compounds. J Agric Food Chem Sep 54: 6547-6553.
- Chen C, Pung D, Leong V, Hebbar V, Shen G, et al. (2004) Induction of detoxifying enzymes by garlic organosulfur compounds through transcription factor Nrf2: effect of chemical structure and stress signals. Free Radic Biol Med 37: 1578-1590.
- Yin X, Zhang R, Feng C, Zhang J, Liu D, et al. (2014) Diallyl disulfide induces G2/M arrest and promotes apoptosis through the p53/p21 and MEK-ERK pathways in human esophageal squamous cell carcinoma. Oncol Rep 32: 1748-1756.
- Shin DY, Kim GY, Lee JH, Choi BT, Yoo YH, et al. (2012) Apoptosis induction of human prostate carcinoma DU145 cells by diallyl disulfide via modulation of JNK and PI3K/AKT signaling pathways. Int J Mol Sci 13: 14158-14171.
- Singh SV, Mohan RR, Agarwal R, Benson PJ, Hu X, et al. (1996) Novel anti-carcinogenic activity of an organosulfide from garlic: inhibition of H-RAS oncogene transformed tumor growth in vivo by diallyl disulfide is associated with inhibition of p21H-ras processing. Biochem Biophys Res Commun 225: 660-665.
- Druesne-Pecollo N, Latino-Martel P (2011) Modulation of histone acetylation by garlic sulfur compounds. Anticancer Agents Med Chem 11: 254-259.
- Altonsy MO, Habib TN, Andrews SC (2012) Diallyl disulfide-induced apoptosis in a breast-cancer cell line (MCF-7) may be caused by inhibition of histone deacetylation. Nutr Cancer 64: 1251-1260.
- Myzak MC, Dashwood RH (2006) Histone deacetylases as targets for dietary cancer preventive agents: lessons learned with butyrate, diallyl disulfide, and sulforaphane. Curr Drug Targets 7: 443-452.
- Zhou Y, Su J, Shi L, Liao Q, Su Q (2013) DADS downregulates the Rac1-ROCK1/PAK1-LIMK1-ADF/cofilin signaling pathway, inhibiting cell migration and invasion. Oncol Rep 29: 605-612.
- Lea M, Ayyala U (1997) Differentiating and growth inhibitory effects of diallyl disulfide on cancer cells. Int J Oncol 11: 181-185.
- Hahm ER, Singh SV (2014) Diallyl trisulfide inhibits estrogen receptor-alpha activity in human breast cancer cells. Breast Cancer Res Treat 144: 47-57.
- Lei XY, Yao SQ, Zu XY, Huang ZX, Liu LJ, et al. (2008) Apoptosis induced by diallyl disulfide in human breast cancer cell line MCF-7. Acta Pharmacol Sin 29: 1233-1239.
- Nagaraj NS, Anilakumar KR, Singh OV (2010) Diallyl disulfide causes caspase-dependent apoptosis in human cancer cells through a Bax-triggered mitochondrial pathway. J Nutr Biochem 21: 405-412.
- Xiao X, Chen B, Liu X, Liu P, Zheng G, et al. (2014) Diallyl disulfide suppresses SRC/Ras/ERK signaling-mediated proliferation and metastasis in human breast cancer by up-regulating miR-34a. PLoS One 9: e112720.
- Rathod SS, Rani SB, Khan M, Muzumdar D, Shiras A (2014) Tumor suppressive miRNA-34a suppresses cell proliferation and tumor growth of glioma stem cells by targeting Akt and Wnt signaling pathways. FEBS Open Bio 4: 485-495.
- Li XJ, Ren ZJ, Tang JH (2014) MicroRNA-34a: a potential therapeutic target in human cancer. Cell Death Dis 5: e1327.
- Na HK, Kim EH, Choi MA, Park JM, Kim DH, et al. (2012) Diallyl trisulfide induces apoptosis in human breast cancer cells through ROS-mediated activation of JNK and AP-1. Biochem Pharmacol 84: 1241-1250.
- Bauer D, Redmon N, Mazzio E, Taka E, Reuben JS, et al (2015) Diallyl disulfide inhibits TNFalpha induced CCL2 release through MAPK/ERK and NF-Kappa-B signaling. Cytokine 75: 1-10.
- Bauer D, Mazzio E, Soliman KF, Taka E, Oriaku E, et al. (2014) Diallyl disulfide inhibits TNFalpha-induced CCL2 release by MDA-MB-231 cells. Anticancer Res 34: 2763-2770.
- Arunkumar A, Vijayababu MR, Kanagaraj P, Balasubramanian K, Aruldhas MM, et al. (2005) Growth suppressing effect of garlic compound diallyl disulfide on prostate cancer cell line (PC-3) in vitro. Biol Pharm Bull 28: 740-743.
- Arunkumar A, Vijayababu MR, Srinivasan N, Aruldhas MM, Arunakaran J. (2006) Garlic compound, diallyl disulfide induces cell cycle arrest in prostate cancer cell line PC-3. Mol Cell Biochem 288: 107-113.
- Xiao D, Choi S, Johnson DE, Vogel VG, Johnson CS, et al. (2004) Diallyl trisulfide-induced apoptosis in human prostate cancer cells involves c-Jun N-terminal kinase and extracellular-signal regulated kinase-mediated phosphorylation of Bcl-2. Oncogene 23: 5594-5606.
- Ruvolo PP, Deng X, May WS (2001) Phosphorylation of Bcl2 and regulation of apoptosis. Leukemia 15: 515-522.
- Toren P, Zoubeidi A (2014) Targeting the PI3K/Akt pathway in prostate cancer: challenges and opportunities (review). Int J Oncol 45: 1793-1801.
- Arunkumar R, Sharmila G, Elumalai P, Senthilkumar K, Banudevi S, et al. (2012) Effect of diallyl disulfide on insulin-like growth factor signaling molecules involved in cell survival and proliferation of human prostate cancer cells in vitro and in silico approach through docking analysis. Phytomedicine 19: 912-923.
- Arunkumar A, Vijayababu MR, Gunadharini N, Krishnamoorthy G, Arunakaran J (2007) Induction of apoptosis and histone hyperacetylation by diallyl disulfide in prostate cancer cell line PC-3. Cancer Lett 251: 59-67.
- Chen WC, Hsu SS, Chou CT, Kuo CC, Huang JK, et al. (2011) Effect of diallyl disulfide on Ca2+ movement and viability in PC3 human prostate cancer cells. Toxicol In Vitro 25: 636-643.
- Rahim F, Hajizamani S, Mortaz E, Ahmadzadeh A, Shahjahani M, et al. (2014) Molecular regulation of bone marrow metastasis in prostate and breast cancer. Bone Marrow Res 2014: 405920.
- Steeg PS, Theodorescu D (2008) Metastasis: a therapeutic target for cancer. Nat Clin Pract Oncol 5: 206-219.
- Shin DY, Kim GY, Kim JI, Yoon MK, Kwon TK, et al. (2010) Anti-invasive activity of diallyl disulfide through tightening of tight junctions and inhibition of matrix metalloproteinase activities in LNCaP prostate cancer cells. Toxicol In Vitro 24: 1569-1576.
- Lai KC, Kuo CL, Ho HC, Yang JS, Ma CY, et al. (2012) Diallyl sulfide, diallyl disulfide and diallyl trisulfide affect drug resistant gene expression in colo 205 human colon cancer cells in vitro and in vivo. Phytomedicine 19: 625-630.
- Germain E, Auger J, Ginies C, Siess MH, Teyssier C (2002) In vivo metabolism of diallyl disulphide in the rat: identification of two new metabolites. Xenobiotica 32: 1127-1138.