Research Article
Effect of Maternal Protein Restriction during E10&11 Day of Gestation on Organ Weights, Litter Size, Placental Weight and Biochemical Analysis
A Benno Susai Vijayakumar1*, M Patrick Gomez2
1Department of Biochemistry, St. Joseph’s College (Autonomous) Tiruchirappalli, 620 002, Tamil Nadu, India
2Department of Botany, Biochemistry and Biotechnology, St. Joseph’s College (Autonomous) Tiruchirappalli, 620 002, Tamil Nadu, India
Corresponding author: A Benno Susai Vijayakumar, Department of Biochemistry, St.Joseph’s College (Autonomous)Tiruchirappalli, 620 002, Tamil Nadu, India; E-mail: bennosusai@gmail.com
Citation: Longkumer T. Age and sex differences in human body physique and its association with nutrition: A cross-sectional study among the A children from Nagaland, North-East India. Indian J Nutri. 2016;3(2): 132.
Copyright © 2016 Longkumer T. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Indian Journal of Nutrition | ISSN: 2395-2326 | Volume: 3, Issue: 2
Submission: Submission: 16/07/2016; Accepted: 30/07/2016; Published: 05/08/2016
Abstract
Environmental constraints during pregnancy results in phenotypic changes which will be associated with changes in biochemical components and alteration in body and placentae weight. The aim of the study is to investigate the effect of maternal protein restriction during embryonic day 10 (E10) and 11 (E11) (Acute protein restriction). Pregnant mice were divided into four groups (I-IV) respectively. Pregnant mice were fed on 10th and 11th day with isocaloric low protein diets (control diet 24% diet for group I, 18% diet for group II, 12% diet for group III and 6% diet for group IV) and studied at gestational day 18. Maternal body weight, visceral organ and reproductive organ weights, litter size, placental number and biochemical parameters were analyzed. The levels of various biochemical parameters including serum glucose, urea, uric acid, creatinine, cholesterol, HDL, LDL, TGL, VLDL, bilirubin (total, indirect and direct) showed significant variations (PË‚0.05) when compared to those of the control group. Similarly, SGPT, SGOT, total protein and albumin levels increased significantly in protein restricted groups, when compared with group I, whereas the body weight and organ weights showed no significant change. Among the restricted groups,group IV shows 5.0771 ± 0.128 g, group III 5.439 ± 0.059 g, group II 55.2501 ± 0.026 g and control group I with 5.5056±0.0341 g of placenta weights. While the weights of fetuses in group I26.81 ±2.993, group II 13.76 ±0.211, group III 18.3±2.151 and group IV 16.16 ±1.970 g shows reduction in placental weights. We demonstrated the effect of maternal protein restriction during 10 and 11th day of gestation, which influences the change in the visceral organ weight and reproductive organ weight. Our observation coincides with the earlier findings which showed that the increase in the blood glucose in the restricted group paved way for the development of Diabetes in the adult life and showed lower fetal weight and placental weight in the restricted groups.
Keywords: Placentae; Maternal protein restriction; Acute protein restriction; IUGR; Diabetes
Introduction
Introduction of the fetus to maternal malnutrition is wellrecognized causal factor for intrauterine growth restriction (IUGR),in both humans and animals. Low protein diets in particular havebeen related with distorted placental growth and composition, fetalgrowth and organ development [1-4].
Maternal nutrition during gestation has a pivotal role in theregulation of placental-fetal development and in that way affects theall-time health and productivity of the offspring in human and animalmodels [5]. The maternal suboptimal nourishment during pregnancyresults in IUGR and newborns with low birth weight. Intrauterinegrowth restriction is associated with increased perinatal morbidityand death, and newborns with low birth weight have increased the threat for the development of adult metabolic syndrome [6].Maternal protein restriction adversely affects nutrient supply to thefetus. Human epidemiological observations from the ‘Dutch HungerWinter’ studies have revealed that adults born to mothers exposed topoor nutrition during early pregnancy displayed increased early-onsetincidences of coronary artery diseases [7]. Maternal malnutrition,psychological stress or perturbed hormonal status transmit signalsto the developing fetus and, depending on the timing of theseexposures, permanently changing tissue structure and function[8]. In 1970’s, Forsdahl, using official data on Norwegian counties,reported that scarcity during adolescence, followed by prosperity,was positively correlated with risk of death from coronary heartdiseases [9]. In 1986, Barker and colleagues began publishing reportson the association between an adverse intrauterine environment, asdetermined primarily by low birth weight, and an increased risk ofcoronary heart disease later in life [10]. Collectively, the above studies generated the Forsdahl-Barker hypothesis, recognizing Forsdahl,as the original basis of the idea and Barker as the developer of theconcept. The Forsdahl-Barker hypothesis has become better knownas the “fetal origins” hypothesis and has created a new branch ofscientific knowledge and investigation, known as “The DevelopmentalOrigins of Health and Diseases”(DOHaD) [11]. It is now broadlyacknowledged that certain chronic diseases of adulthood may havetheir genesis in the womb. From the above literature, we hypothesizedthat maternal protein restriction from E10 and E11of gestation (acuteprotein restriction), in mouse, would result in changes in litter size,placental weight, and organ weight and variations in biochemicalparameters of maternal serum.
Materials and Methods
Animals and experimental treatments
The experimental study was carried out using weaning Swissalbino mice weighing about 15-20 grams. The animals were housedindividually and fed with control or normal diet, till conception adlibitum. Female mice were housed with males, of proven fertilityfor mating. Next morning, the vaginal plug positive animals wereseparated (n6) and grouped into four. Group I- IV animals were fedwith normal control diet (100% protein diet). On 10th and 11th dayof gestation, animals in groups II, III and IV were fed with proteinrestricted diet (25% protein restricted diet, 50% protein restricteddiet and 75% protein restricted diet) respectively (Table 1). The micewere fed with modified Laboratory Animal Information ServiceCentre diet. The experimental design was approved by InstitutionalAnimal Ethical Committee, St.Joseph’s College (Autonomous),Tiruchirappalli.
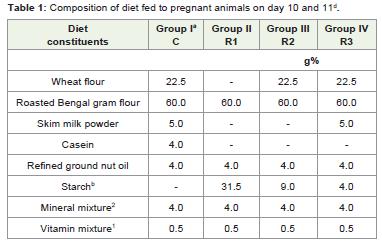
Table 1: Composition of diet fed to pregnant animals on day 10 and 11d.
Collection of tissue and serum sample
At day 18, animals were euthanized (Group -I, II, III, & IV with6 animals each) with ether. After dissection, serum, organs, foetus,placentae were collected quickly. The liver, kidney, heart, spleen,placentae were dried on blotting paper and their weights weremeasured. The maternal serum was stored at -20 °C for biochemicalestimations.
Determination of serum biochemical parameters: Serum fromthe respective groups was pooled for various biochemical parameters.
All the parameters were analysed with Prietest Clinical Biochemistryreagents kits using Prietest Touch-Auto Biochemistry Analyser,(Robonik India Pvt, Ltd.).
Determination of body weight and organ weights: The bodyweights of all the groups were measured once in three days fromconception till sacrifice. The organ weights, fetal number, and theplacentae number of the respective groups were measured.
Statistical analysis
Data are presented as means ± SEM. Statistical analyses wereperformed using student’s t-test, where significant differences wereindicated. A probability of 5% or less was accepted as significant.
Result
In this study, a total of 24 female pregnant animals were studied. The growth increment of the mother, foetus and the major organsin respective groups were measured. The maternal weights of therespective groups increase with the gestational age (Table 2). Thereduction in the protein content of the diet did significantly affectthe weight of the dams at slaughter on day 18.The number of foetusescarried by each mother were dissimilar in mice given normal dietsafter protein restriction. While the weight of the placenta in restrictedgroups show significant reduction, the weights of the foetuses ingroup 3 are increased than those of the group II and IV, though thefetal number in group III is less as compared to restricted group II(Table 3).
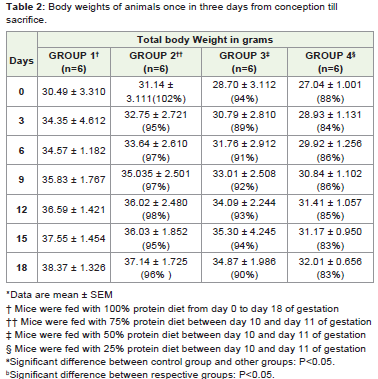
Table 2: Body weights of animals once in three days from conception till sacrifice.
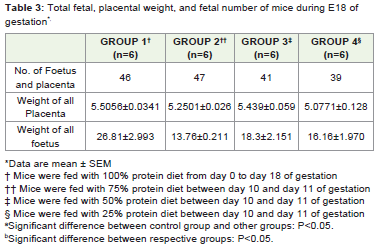
Table 3: Total fetal, placental weight, and fetal number of mice during E18 of gestation*.
A significant (p< 0.05) increase in serum glucose levels wereobserved in control group (Group I) animals as compared to thosefrom other three groups (Group II, III & IV). Among the groups, the highest concentration of serum glucose level was recorded in proteinrestricted group IV 101.50 ± 1.76 mg/dL, which is 122% increaseas compared to control group I 83.00 ± 1.41mg/d/L, followed bygroup II 90.00 ± 1.41mg/dLwith 108% increase and group III 80.83± 0.76 mg/dL with 92% increase (Table 4). A significant (P0.05) raise in serum urea level was noted in group III50.00 ± 0.90mg/dL and group IV54.83 ± 0.75mg/dL as compared to group I44.33 ± 1.63mg/dL. Creatinine levels in the restricted group III 2.71 ± 0.15mg/dLincreases significantly as compared to the control group I 2.38± 0.17mg/dL. Increase in cholesterol levels were also noticed in theprotein restricted groups (Group III 80.66 ± 0.82; 101% and Group IV87.33 ± 0.82mg/dL; 109%) as compared to the control group I79.67± 1.03mg/dL; 100%. In case of HDL and LDL cholesterol levels, theserum concentration of the above two were increased significantly inthe group IV when compared to the control group.
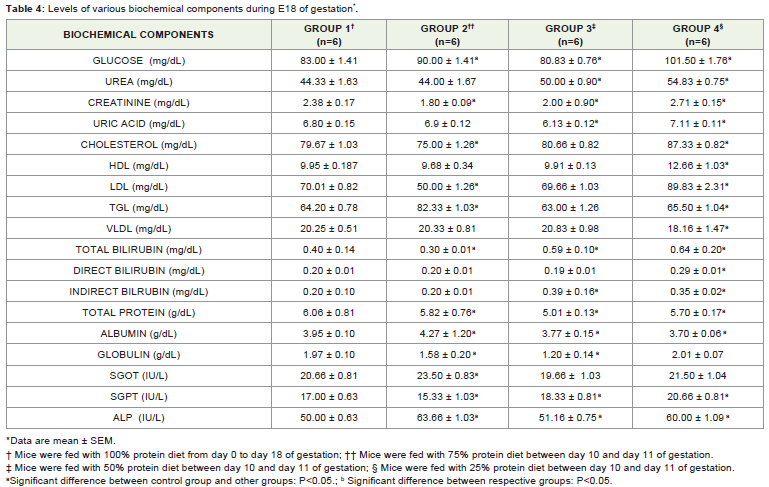
Table 4: Levels of various biochemical components during E18 of gestation*.
The levels of HDL and LDL were decreased in group III whencompared to the group IV. The serum VLDL levels in the respective groups show significant decrease in the restricted group IV 18.16 ±1.47mg/dL; 89% as compared to the control group I 20.25 ± 0.5mg/dL; 100%. In comparison with control group I 64.20 ± 0.78mg/dL; 100%, TGL is elevated 128% in group II 82.33 ± 1.03mg/dL ascompared to group III 63.00 ± 1.26 mg/dL and group IV 65.50 ± 1.04mg/dL. No significant change is observed in group IV (102%).
Serum bilirubin (Total, direct and indirect) was significantlyincreased in the restricted groups II, III & IV when compared to thatof the control group. In protein metabolism, the utilization of thetissue/serum proteins in the restricted groups results in the decreaseof the total protein and albumin levels when compared to the controlgroup.
Serum enzyme levels were significantly increased in SGPT andALP. The level of SGOT in group 2 increased (13%) when comparedto that of the other restricted groups.
Organ weights in the restricted group IV showed significantincrease. The percentage of increase in the weights of liver 2.14 ±0.21g; spleen 0.20 ± 0.05g; and, uterus and ovary 0.84± 0.20 g; are138%, 166% and 116% respectively. While the weight of heart in therestricted group remains the same, the weight of the kidney in therestricted group IV shows a slight decrease (3%) and the weight ingroup II was significantly reduced (13%) (Table 5).
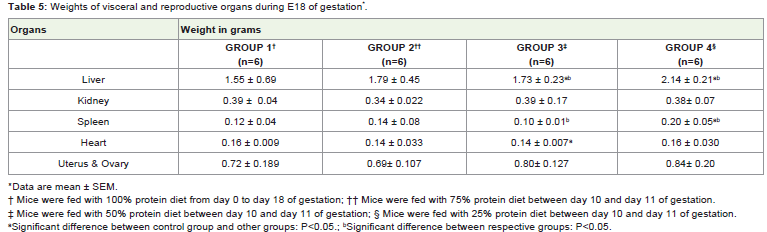
Table 5: Weights of visceral and reproductive organs during E18 of gestation*.
Discussion
The present study addressed the effects of maternal proteinrestriction mediated maternal stress during E10 and E11 day ofgestation on fetal body weight, organ weight, placental number andbiochemical parameters. Maternal protein restriction produced apredictable stress response in the mother, as evidenced by elevatedmaternal plasma glucose levels in the restricted groups. Our resultsshowed decrease in the fetal weight and placenta in the E10 and E11 day of the pregnancy in mice. Langley-Evans et al. (1996) andGonzalez et al., observed a similar early increase in the growth of thefoetuses in pregnant rats of the Wistar strain [12,13].
Numerous animal studies have investigated the effect of maternallow-protein diet on glucose metabolism in the off-spring [14]. Wehypothesized the impact of maternal low protein diet during E10and E11 day of gestation in mice, which shows significant increasein serum glucose in the protein restricted groups (Group II and IV).Similarly, a low protein diet during gestation (10% protein in therestricted group) shows no significant difference in serum glucoseduring E19 in rats [15]. The levels of TGL show no significant changein previous study by Nimbe Torres et al., (2010) [16]. We previouslyreported that maternal protein restriction determines the fate anddevelopment of the foetus during development and the origin of adultdisease in later life [17].
Conclusion
Concept and importance of nutritional programming wasrecommended by epidemiological studies in human duringpregnancy and confirmed by animal experiments (Swine, Sheep, Ratand Mice). In humans, as it was proved by numerous papers, scientificfield of nutritional programming is hopeful and represents a theme of major public health and clinical significance. In this perspective,it is necessary to define defensive and predisposing effects of earlynutrition on the development of later chronic diseases, since earlynutrient feeding can be potentially modified to minimize the risk ofdiseases in later life. In conclusion, our study also showed that thedietary protein restriction during mid gestation in mouse (Acuteprotein restriction) resulted in decreased fetal and placental weight asevident from various works. Our observation coincides with the earlyfindings of increased blood glucose in the restricted group paved theway for the development of Diabetes during adult life. Further insightis essential in the Developmental Origins of Health and Diseases(DOHaD) in later life and the involvement of placenta or placental gene expression of the same.
Acknowledgment
Authors sincerely thank Prof. Antony Diwakar Chandran,Assistant Professor, Department of Biochemistry, St.Joseph’sCollege (Autonomous), Tiruchirappalli for his valuable technicalassistance during manuscript preparation, Dr. Karthik Mohan,Assistant Professor, Department of Biochemistry, St.Joseph’s College(Autonomous), Tiruchirappalli for his expertise and proof readingand, Dr. B. Chitra, Assistant Professor, Department of Biotechnology,Srimad Andavan Arts and Science College (Autonomous), Tiruchirappalli for her assistance.
References
- Ariyuki F (1987) Growth retardation induced in rat fetuses by maternal fasting and massive doses of ergocalciferol. J Nutr 117: 342-8.
- Mello MAR, Cury L, Valle LBS and Oliveira-Filho RM (1987) Protein-calorie malnutrition in the young pregnant rat: factors involved in fetal growth impairment. Bra J Med Biol Res 20: 575-7.
- Godfrey KM, Redman CWG, Barker DJP, Osmond C (1991) The effect of maternal anaemia and iron deficiency on the ratio of fetal weight to placental weight. Br J Obstet Gynaecol I: 866-91.
- Mellor DJ (1983) Nutritional and placental determinants of foetal growth rate in sheep and consequences for the newborn lamb. Br Vet J 139: 307-24.
- Belkacemi L, Nelson DM, Desai M, Ross MG (2010) Maternal undernutrition Influences Placental-Fetal Development. Biology of reproduction 83: 325-331.
- Marsal K (2002) Intrauterine growth restriction. Curr Opin Obstet Gynecol 14: 127-135.
- Barker DJP, Clark PM (1997) Fetal undernutrition and disease in later life. Rev Reprod 2: 105-112.
- Painter RC, de Rooij SR, Bossuyt PM, Simmers TA, Osmond C, et al. (2006) Early onset of coronary artery disease after prenatal exposure to the Dutch famine. Am J Clin Nutr 84: 322-327.
- Hales CN, Barker DJ (1992) Type 2 (non-insulin-dependent) diabetes mellitus: the thrifty phenotype hypothesis. Diabetologia 35: 595-601.
- Forsdahl A (1977) “Are poor living conditions in childhood and adolescence an important risk factor for arteriosclerotic heart disease?” Br J Prev Soc Med 31: 91-95.
- Barker DJ, Osmond C (1988) “Low birth weight and hypertension,” British Medical Journal 297: 134-135.
- Vangen S, Nordhagen R, Lie KK (2005) Revisiting the Forsdahl-Barker hypothesis. Journal of the Norwegian Medical Association vol. 125: 451-453.
- Langley-Evans SC, Gardner DS, Jackson AA (1996) Association of disproportionate growth of fetal rats with raised systolic blood pressure in later life. J Reprod Fertil 106: 307-312.
- Gonzalez PN, Gasperowicz M, Barbeito-Andrés J, Klenin N, Cross JC, et al. (2016) Chronic protein restriction in mice impacts placental function and maternal body weight before fetal growth. PLoS ONE 11: 1-18.
- Fernandez-Twinn DS, Wayman A, Ekizoglou S, Martin MS, Hales CN, et al. (2005) Maternal protein restriction leads to hyperinsulinemia and reduced insulin-signaling protein expression in 21-mo-old female rat offspring. Am J Physiol Regul Integr Comp Physiol 288: R368-R373.
- Torres N, Bautista CJ, Tovar AR, Ordáz G, RodrÃguez-Cruz M (2010) Protein restriction during pregnancy affects maternal liver lipid metabolism and fetal brain lipid composition in the rat Am J Physiol Endocrinol Metab 298: E270-E277.
- Vijayakumar ABS, Gomez M P (2016) Maternal protein restriction determines the fate of the fetus - a Review. Int J Health Sci Res 6: 341-348.