Differential Responses of Antioxidant Enzymes in Salt-Stressed Almond Tree Grown Under Sun and Shade Conditions
Ahlem Zrig1, Hatem Ben Mohamed2, Taieb Tounekti1,3, Sarra Oueled Ahmed1 and Habib Khemira1,3*
Corresponding author: Dr. Ahlem ZRIG, Research Unit of Biodiversity and Valorization of Bioresources in Arid Zones,Faculty of Sciences of Gabes- City Erriadh, Zrig, Gabes 6072, Tel: (+216) 97 901 249, Tunisia; E-mail: ahlem18zrig@yahoo.fr
1Research Unit of Biodiversity and Valorization of Bioresources in Arid Zones, Faculty of Sciences of Gabes- City Erriadh, Zrig,Gabes 6072, Tunisia
2Laboratory of Horticulture, Tunisian National Agricultural Research Institute (INRAT), Rue Hédi Karray 2049 Ariana,Tunisia
3Centre for Environmental Research & Studies (CERS), Jazan University, Jazan, Kingdom of Saudi Arabia
Citation: Zrig A, Mohamed HB, Tounekti T, Ahmed SO, Khemira H. Differential Responses of Antioxidant Enzymes in Salt-Stressed Almond Tree Grown02 Under Sun and Shade Conditions. J Plant Sci Res. 2015;2(1): 117.
Copyright © 2014 Ahlem Zrig et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Plant Science & Research | ISSN: 2349-2805 | Volume: 1, Issue: 4
Submission: 03/12/2014; Accepted: 29/01/2015; Published: 02/02/2015
Abstract
We studied the effects of solar irradiance on antioxidant enzymes and polyphenols in sweet almond cv. Mazzetto grown in salt soil. Photosynthetic performance, leaf pigments, activities of antioxidant enzymes and concentrations of polyphenols were measured in plants growing in shade or full sun and supplied with 100 mM NaCl over a 4-week period. In response to the salt stress treatment, photosynthetic rates decreased more in sun plants than in shade plants, likely because of declines in chlorophyll and carotenoids content coupled with significant reductions in stomatal conductance and MDA increases. Antioxidant enzymatic activities such as APX and POD were much greatest in sun leaves under unstressed conditions, however it increased in response to salt stress more in shade leaves. Whereas, Cat decreased under all treatments. Polyphenols markedly increased in response to full sunlight irrespective of salt treatment; however, polyphenols concentrations increased more in response to salt stress in shade leaves.
Keywords: Salinity, Antioxidant enzymes, Polyphenols, Gas exchange, Leaf pigments, Solar irradiation, MDA
Introduction
The response of plants to combination of two environmental stresses is unique and cannot be extrapolated from the responses to individual or separate stress. Salinity and High light (HL) intensity are the major environmental stress frequent in arid regions. Under both stress, trees are liable to suffer photoinhibition, defined as the slow, reversible decline in photochemical efficiency that occurs under photooxidative stress [1]. Sunlight contains high-energy ultraviolet radiation (UV, 280–400 nm) and photosynthesis is one of the processes most sensitive to high irradiance [2]. In this case, the generations of reactive oxygen species (ROS) in photosynthetic tissues are enhanced [3]. ROS accumulation is a common event in leaves of stressed plants [4] where they cause various damages to macromolecules and cell structures [5]. Nevertheless, both stresses activate common responses, which mostly operate at the biochemical level, to electively counter the oxidative damage [6]. High sunlight and root-zone salinity may also alter photosystem II (PSII) photochemistry and photosynthetic pigment composition [7], by either decreasing the total chlorophyll (Chl) concentration [8] or increasing the ratio of violaxanthin-cycle pigments to Chltot. These biochemical adjustments effectively reduce the risk of photodamage and enhance the dissipation of excess energy via non-photochemical (NP) processes [9]. Moreover, carotenoids may inhibit the generation of free radicals [10]. Likewise, the high sunlight and salinity stress enhance the biosynthesis of polyphenols, particularly flavonoids, likely using ROS as signalling molecules [11,12]. In turn, the increase in leaf flavonoid content effectively attenuates the penetration of UV wavelengths in sensitive leaf targets [13] and, possibly, counters excess light-induced oxidative damage [14]. In addition, in such conditions, plants developed enzymatic mechanisms involved in the enzymatic scavenging of ROS. These antioxidant mechanisms employ (i) ROS-scavenging enzymes, such as superoxide dismutase (SOD, EC 1.15.1.1), catalase (CAT, EC 1.11.1.6), and ascorbate peroxidase (APX, 1.11.1.11), (ii) recycling enzymes of the ascorbateglutathione cycle, such as monodehydroascorbate reductase (MDHAR, EC 1.6.5.4), dehydroascorbate reductase (DHAR, EC 1.8.5.1) and glutathione reductase (GR, EC 1.6.4.2),and (iii) low molecular weight antioxidants, such as reduced ascorbic acid (Asa) and reduced glutathione (Gsh). Some authors have reported that antioxidative systems play a major role in protecting plants from the harmful effects of excess light energy [15,16]. Thus, antioxidative systems have been found to be of paramount importance in the response and tolerance of trees to environmental stress [16]. The issue of how sunlight irradiance interacts with excess soil salinity in determining a plant’s performance under field conditions, although not much investigated, has great eco-physiological and horticultural relevance [17,11]. Thus, salt stress as rated through ecophysiological metrics is reduced by low light or shading. In this paper, we tend to making one speculate that the interactive effects of light and salinity together allow almond tree to flourish on these soils. Sweet almond cv. Mazzetto cultivated was tested to determine their physiological responses to salinity in sun (High Light) versus shade (Low Light). We hypothesized that high light would intensify the negative effects of salinity on almond growth, and these findings would help weed managers understand how the two stressors can interact to restrict or exacerbate its invasion to croplands. The aim of this study was to characterize the combined effects of salt stress and light conditions on growth, photosynthetic apparatus and the antioxidant systems and to understand the interaction between stress enhancing the polyphenols and the antioxidants enzymes.
Materials and Methods
Plant material and salt treatments
The present study was performed on one year old sweet almond cv Mazzetto. The plants were cultivated in perforated 4L-plastic-pots containing desert dune-sand. They were irrigated every 4 days with a complete nutrient solution (N, 1.8 mM; P, 0.35 mM; K, 0.64 mM; Ca, 1.0 mM; Mg, 0.35 mM; S, 0.35 mM; Fe, 0.03 mM; Zn, 0.4 μM, Mn, 5.0 μM; Cu, 0.1 μM and B, 0.02 mM). The salinity stress was obtained by adding NaCl to the nutrient solution to obtain 25, 50 and 75 mM total ion concentrations. Control treatment consisted of no NaCl added. To avoid osmotic shock, the concentration of the nutrient solution was increased by 25 mM per day till the final salinity level for each treatment was reached. Plants in full sun received a daily dose of 1800 to 2500 μM m-2 s-1 in the PAR. Shade plants were exposed to a daily dose of 500 to 700 μM m-2 s-1 in the PAR. Temperature maxima/ minima were averaged 29/14 °C and 25/19 °C in full sun and shade, respectively. From the moment when the final concentration of NaCl was obtained for the most severe stress level, the concentrations of nutrient solutions were kept constant for the following four weeks. Each plant were harvested in the morning (between 9 to 11 a.m. local time), weighed and divided into two batches; one was frozen in liquid nitrogen and stored at -80 °C for biochemical analyses; the other was washed in de-ionized water, dried at 80 °C in a forced-air oven for 48 hours and ground into a fine powder to pass through a 30-mesh screen for ion analyses.
Growth
Before the treatments were imposed, the tip of main the shootof each plant was marked to be able later to assess shoot extension during the period of the experiment. After four weeks of applying saline water and light treatment, the plants were harvested. The latter were washed free of soil particles. The material was then dried for 48h at 80 °C and total dry weights were determined.
Ion content and selectivity
For ion analyses, 20 mg of dry ground leaves from each plantwas extracted with 20 mL of 0.1M HNO3. After filtration, Na+, Ca2+, Mg2+ and K+ contents were determined with an atomic absorption spectrometer (Avanta, GBC, Australia).
Gas exchange measurements
Gas exchange measurements were carried out after 30 days ofsalt treatment. Net photosynthetic rate (A), transpiration rate (E) and stomatal conductance (Gs) of upper leaves were measured with a portable photosynthesis system (Lcp pro+, ADC Systems Ltd, UK)under ambient conditions.
Determination of chlorophylls and carotenoids
Leaf chlorophyll and carotenoids contents were determined byusing the method of Arno (1949) [18]. In short, 0.5 g of fresh leaves were ground to a fine powder in liquid nitrogen in a pre-cooled mortar and homogenized for 30 s in 5 ml of 95.5% acetone. The pigments’ concentrations were estimated from absorbance at 647 nm and 664 nm. A solution of 95.5% acetone was used as a blank. Pigment concentrations were calculated as follows: Chl a (mg/g FW) = [12.7× (A664) _ 2.69× (A647)] × (0.5×5), Chl b (mg/g FW) = [22.9×(A647) _ 4.69× (A664)] × (0.5×5).
Total carotenoids were extracted in duplicates according toMinguez-Mosquera and Hornero-Mendez (1993) [19]. One gram of frozen leaf tissue was briefly extracted with acetone and shaken with diethyl ether and 10% NaCl. Two phases were obtained; the lipophilic phase was washed with Na2SO4 (2%), saponified with 10% KOH in MeOH, and the pigments were subsequently extracted with diethyl ether, evaporated and then made up to 25 ml with acetone. Total carotenoids were estimated by reading the absorbance at 450nm in a UNICAM Helios- spectrophotometer (Cambridge, UK), and expressed as mg of β-carotene equivalent per Kg fresh weight, taking into account the molar absorption coefficient (ε1% cm) of 2560. The results are presented as means± SE.
Phenolic compound contents
Phenolic compounds were extracted 0.5 g of fresh leaves according to Tomas-Barberan et al. (2001) [20] using water: MeOH(2:8) containing 2mM NaF and quantified using the Foline-Ciocalteu reagent. The results were expressed as mg gallic acid equivalent per kg fresh weight of duplicate determinations made on each subsample
Protein contents
For total protein determination, fresh leaves (0.5 g) were ground with 10 mL of 50 mM cooled K-phosphate buffer (pH 7.8) placed in an ice bath. The homogenate was centrifuged at 6,0009g for 20 min at 4C and the soluble proteins content of the extract was determined by the Coomassie blue dye binding method, using bovine serum albumin as a standard [21].
Lipid peroxidation
The level of lipid peroxidation was determined in terms ofthiobarbituric acid-reactive substances (TBARS) concentration asdescribed by Hodges et al. (1999) [22] with some modifications. Leaf samples of 0.5 g were homogenized in 4 mL of 1% (w/v) trichloroacetic acid (TCA), and then centrifuged at 10,0009g for 10 min. To a 1.5 ml aliquot of the supernatant, 1.5 mL of 20% (w/v) TCA containing 0.5% (w/v) TBA was added. The mixture was heated at 95C for 30 min and then quickly cooled in an ice bath. After centrifugation at 10,0009g for 5 min, the values of absorbance measured at 532, 600 and 440 nm of the supernatant were recorded. The value for aspecific absorption at 600 nm was subtracted and a standard curve of sucrose (from 2.5 to 10 lmol mL-1) was used to correct the results from the interference of soluble sugars in samples. The concentration of TBARS wascalculated using an absorption coefficient of 156 mmol-1cm-1 andthe results expressed as MDA equivalents (MDA).
Enzyme extraction and antioxidant activity assay
Tissue extraction of the samples were prepared for the analyses by homogenizing 1 g of fresh leaf material in 4 mL of ice cold 50 mM K-phosphate buffer (pH 7.0) containing 2 mM Na-EDTA and 1% (w/v) polyvinyl– polypirrolidone (PVPP). The homogenate was centrifuged at 10,0009g at 4 C for 10 min. The supernatant was used for determining the activities of ascorbate peroxydas (AXP),peroxidase (POD) and catalase (CAT).
Ascorbate peroxidase (APx; EC 1.11.1.11) activity wasdetermined by following the decrease in A (extinction coefficient 2.8 mM1cm1290) for 30 s in 1 ml of a reaction mixture containing 50 mM potassium phosphate buffer (pH 7.0), 0.5 mM ASC, and 20 μl of enzyme extract. The reaction was started by enzyme extract. Correction was done for the low, non-enzymatic oxidation of ASC by 0.1 mM H2O2 [23].
Activities of CAT and POD were measured according toOsswald et al. (1992) [24], with some modifications. For CAT, the decomposition of H2O2 was followed by the decline in absorbance at 240 nm. The reaction was initiated by adding enzyme extract to 3 mL of reaction mixture containing 50 mM K-phosphate buffer (pH 7.0), 10 mM H2O2 and 50 μL of enzyme extract. CAT activity was determined as the consumption of H2O2 at 240 nm over a 2-min interval. For POD, the activity was determined by measuring the H2O2 oxidation of guaiacol in the presence of H2O2, and following an increase in absorbance at 470 nm over a 2-min interval. The assay mixture (3 mL) contained 0.33 mM guaiacol, 10 mM K-phosphate buffer (pH 7.0), and 50 μL of enzyme extract. The reaction was initiated by adding 40 mM H2O2. The activity of each antioxidant enzyme was expressed on protein basis.
Results
Growth
At the final harvest, leaf biomass was systematically lower in HL plants as compared to control (LL) plants (Table 1). However, all NaCl treated, including those treated with HL, total dry weight and shoot extension decreased while the root/shoot ratio increased (Figure 1).
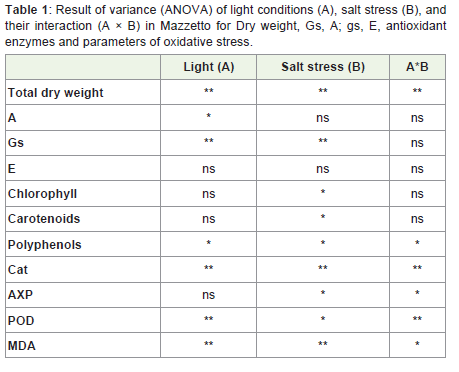
Table 1: Result of variance (ANOVA) of light conditions (A), salt stress (B), andtheir interaction (A × B) in Mazzetto for Dry weight, Gs, A; gs, E, antioxidant enzymes and parameters of oxidative stress.
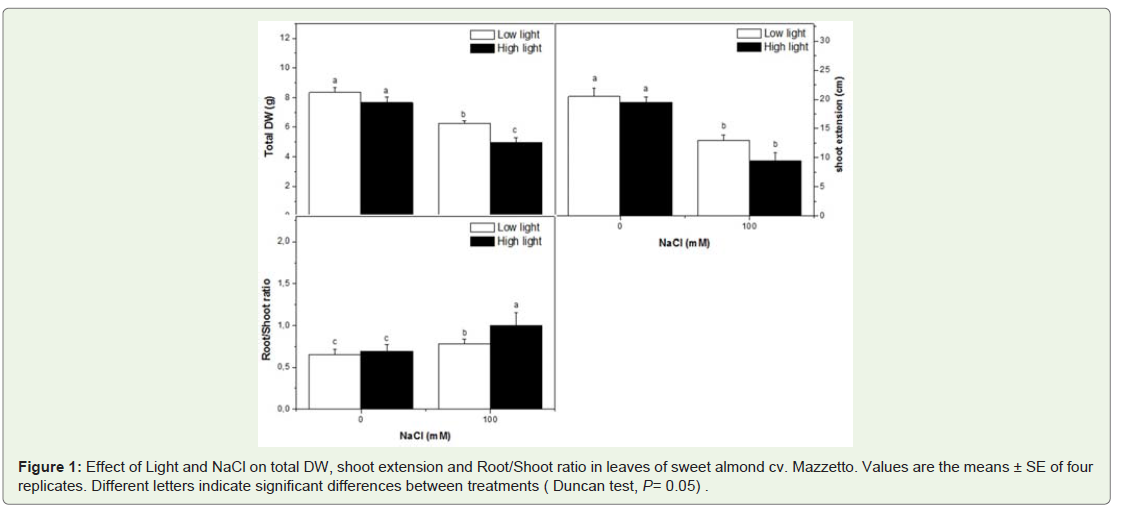
Figure 1: Effect of Light and NaCl on total DW, shoot extension and Root/Shoot ratio in leaves of sweet almond cv. Mazzetto. Values are the means ± SE of four replicates. Different letters indicate significant differences between treatments ( Duncan test, P= 0.05) .
Ion content
Figure 2 shows the mineral contents of leaves as related to the NaCl supply and light treatments. Leaves and roots of salinized almond had higher Na+ concentration than control plants in HL and LL similarly. The effect of HL appeared different on K+ and Mg2+ concentration. The most marked effect presented here is that the higher light intensity resulted in an increased Ca2+ leaf and roots content in salt-stressed plants. In fact, the NaCl treatment depressed significantly the leaf and root K+ and Mg2+ concentration in HL plants more than those in LL. Interestingly, the HL decreased also the leaf and root content of K+ and Mg2+ as comparer to control plant (LL).
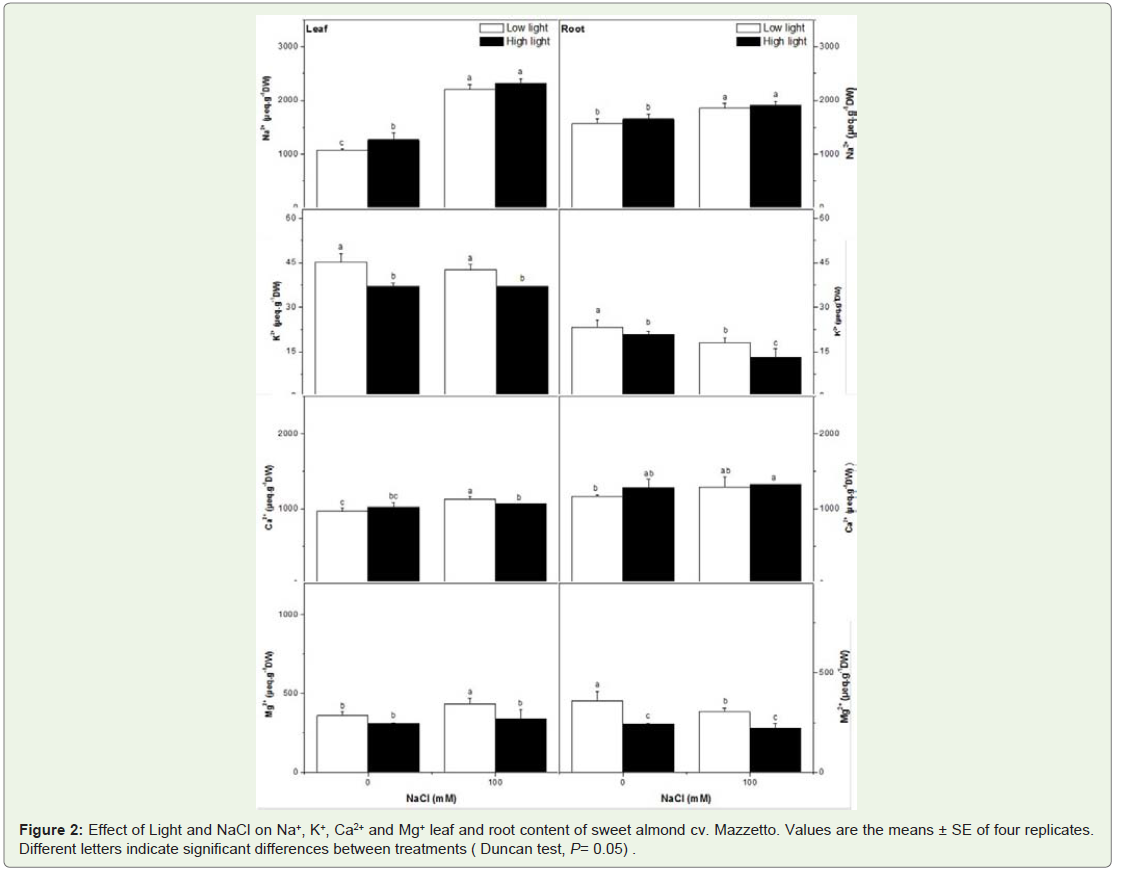
Figure 2: Effect of Light and NaCl on Na+, K+, Ca2+ and Mg+ leaf and root content of sweet almond cv. Mazzetto. Values are the means ± SE of four replicates. Different letters indicate significant differences between treatments ( Duncan test, P= 0.05) .
Gaz exchange
The A decreased in HL plants (10%) as compared to control in response to high light (Figure 3). The slopes of net photosynthetic rate (A) appeared greater for the HL plants than for HL + NaCl stressed leaves. Stomatal conductance (Gs) increased with High Light by 27% as comparer to control plants, and this reduction was greater in NaCl treatment by 90%. Similarly, the net transpiration (E) was more severely depressed in HL plants treated by salinity than untreated plants by NaCl.
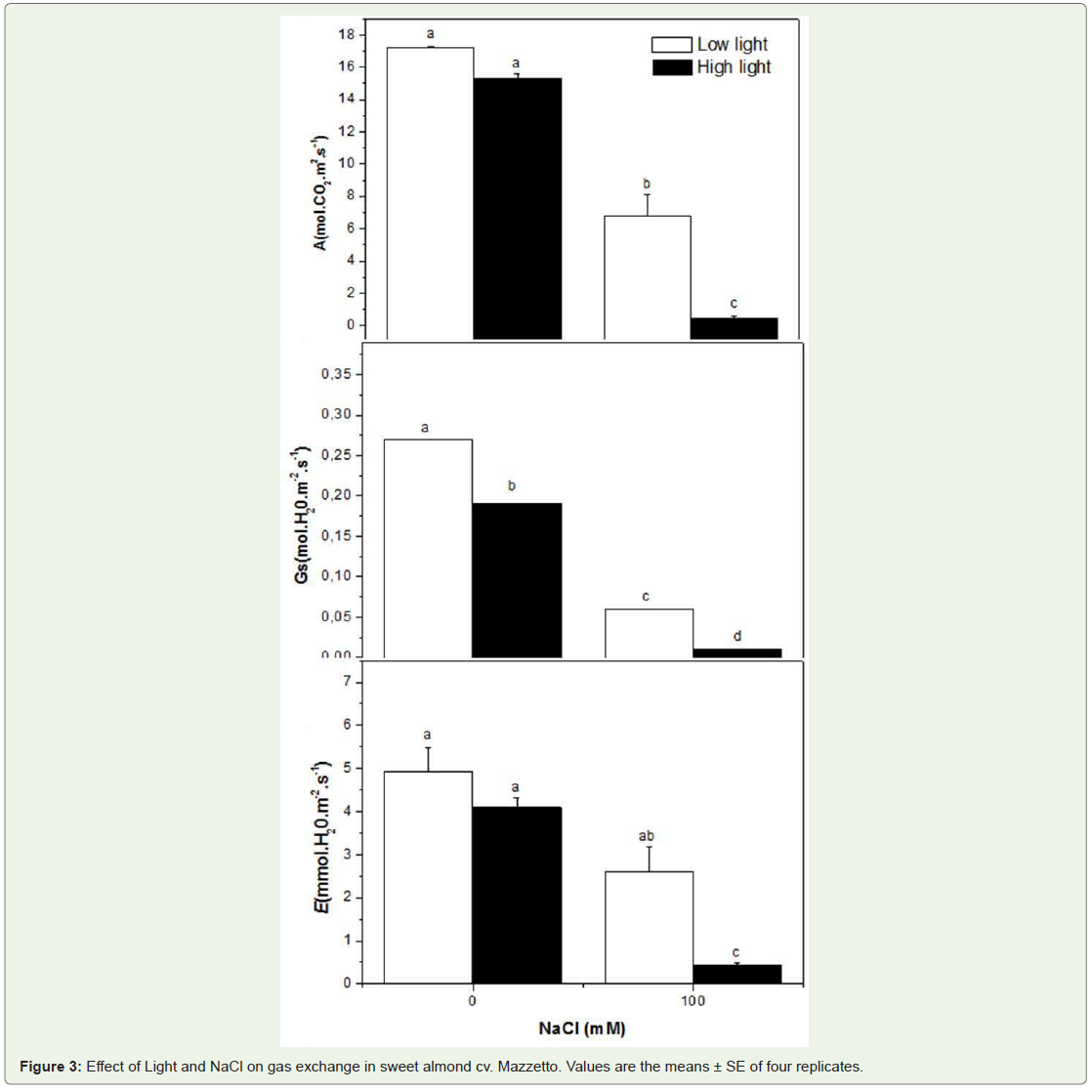
Figure 3: Effect of Light and NaCl on gas exchange in sweet almond cv. Mazzetto. Values are the means ± SE of four replicates.
Chlorophylls and carotenoids
The contents of total chlorophyll (Chl [a+b]) was dependent only on salt conditions (Table 1). Whereas light was found to have no effect on pigment contents in control plant, but it appeared that the sunlight condition increased the salinity effect on pigment. The Chl a/b increased at 100mM NaCl at LL, but decreased when sunlight increased (Figure 4).
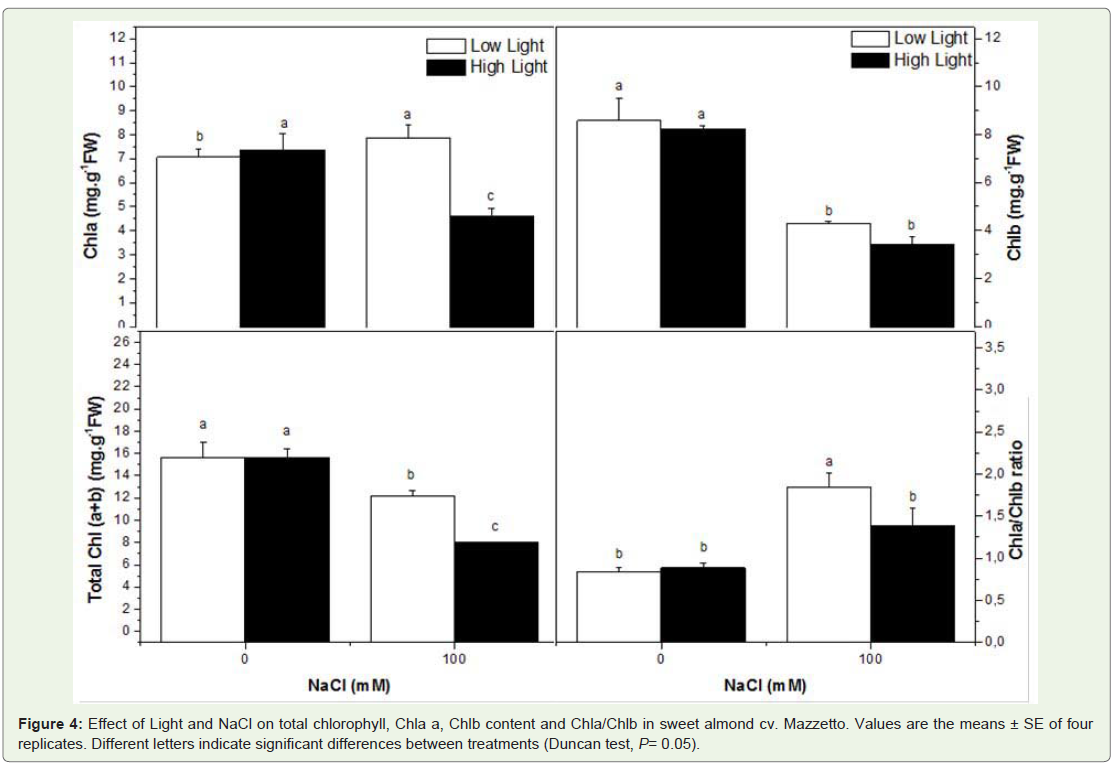
Figure 4: Effect of Light and NaCl on total chlorophyll, Chla a, Chlb content and Chla/Chlb in sweet almond cv. Mazzetto. Values are the means ± SE of four replicates. Different letters indicate significant differences between treatments (Duncan test, P= 0.05).
The concentration and composition of carotenoids varied to a superior degree in response to root-zone salinity than in response to sunlight. In fact, the leaf carotenoids content decrease in all treatment and the higher reduction was recorded in Hl salt stressed plant (48%) comparer to control plant (Figure 5).
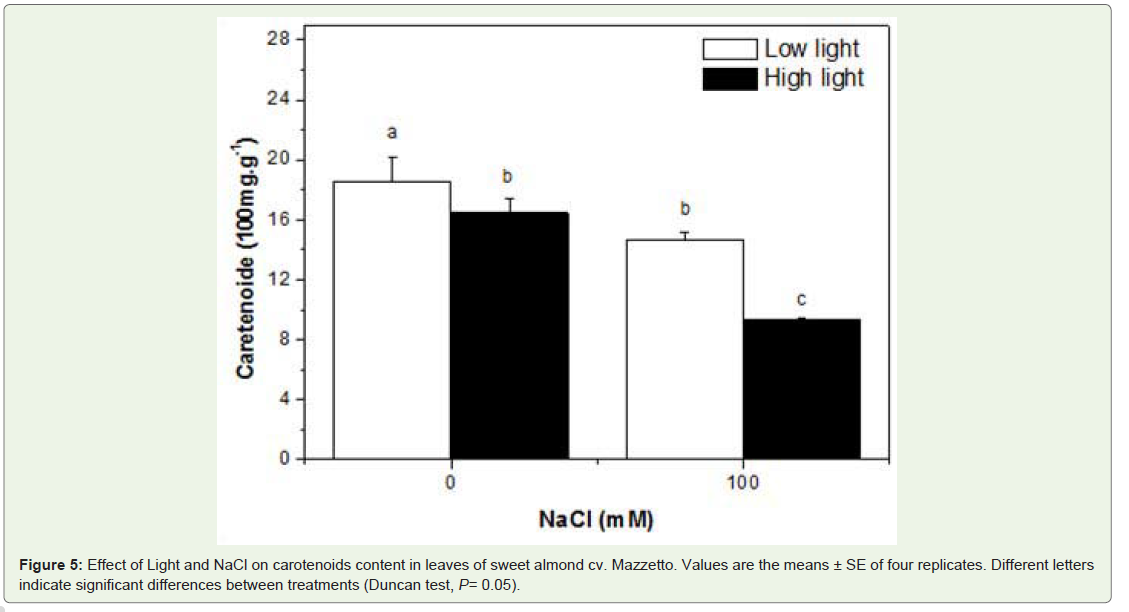
Figure 5: Effect of Light and NaCl on carotenoids content in leaves of sweet almond cv. Mazzetto. Values are the means ± SE of four replicates. Different letters indicate significant differences between treatments (Duncan test, P= 0.05).
Phenolic compound contents
Phenolic leaf content increased with all treatments. The depressive effect of salinity on polyphenols content increased by HL conditions then LL. The highest increase was recorded in HL salt stressed plants as comparer to those in LL (Figure 6).
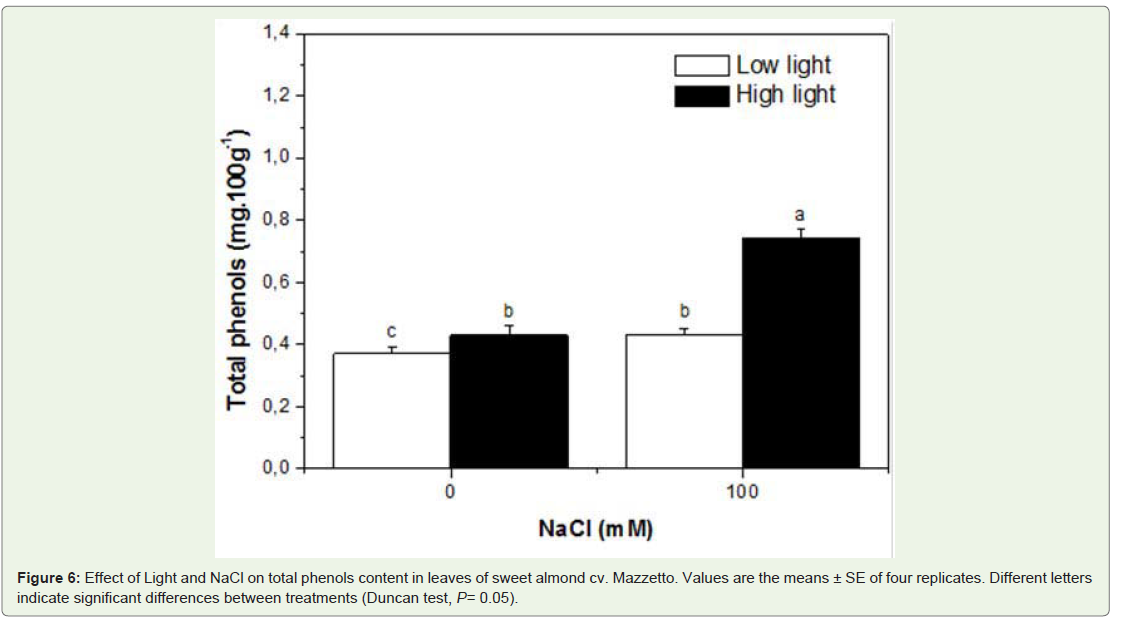
Figure 6: Effect of Light and NaCl on total phenols content in leaves of sweet almond cv. Mazzetto. Values are the means ± SE of four replicates. Different letters indicate significant differences between treatments (Duncan test, P= 0.05).
Lipid peroxidation
To evaluate salt-induced oxidative stress under different light conditions, lipid peroxidation was determined by measurement of MDA content (Figure 7). The MDA content was higher in saltstressed plant treated at LL. Meanwhile, under HL, MDA content decreased in unstressed plants.
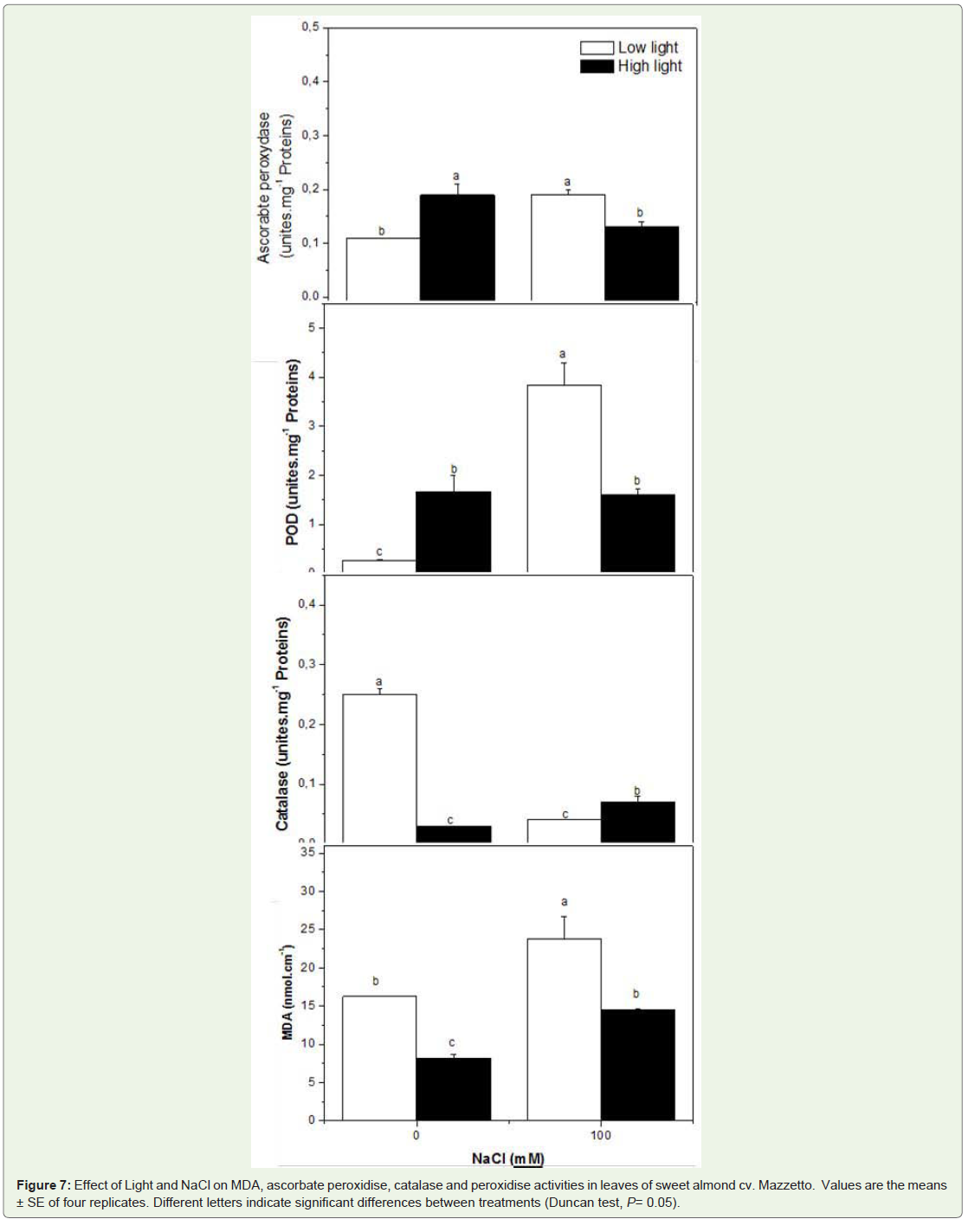
Figure 7: Effect of Light and NaCl on MDA, ascorbate peroxidise, catalase and peroxidise activities in leaves of sweet almond cv. Mazzetto. Values are the means ± SE of four replicates. Different letters indicate significant differences between treatments (Duncan test, P= 0.05).
Antioxidant activity assay
All treatments and their interactions significantly affect the levels of the antioxidants enzymes (Figure 7, Table 1). In all plants, the level of catalase decreased by salt stress under LL or HL. In contrast to the observations of the catalase content, differences in salt-induced changes in the APX and POX antioxidant enzyme activities were observed under different light conditions.
The general trend of ascorbate peroxydase activity shown in Figure 7 suggests that salinity conditions induce higher APX activity. Compared to LL conditions, HL conditions were found to increase the activity of APX by 41%. In contrast, the activity of APX was founded to be reduced by high salinity combined with high irradiation comparer with those treated with HL or salinity alone. In plants under LL, levels of POX was markedly increased in salt stressed plants by 14-fold. While, the POD activity increase in plants in HL and in by 83% and 82%.
Total soluble proteins
Total soluble proteins in the leaves of Mazzetto varied more in response to salinity then to the light treatment. The total protein increased by 15% in response to full sun interactive with salinity (Figure 7).
Discussion
In this experiment, we investigated the effects of salt stress and solar irradiance on the performance of sweet almond cv. Mazzetto to determine the extent of their interactions on biochemical traits, which, in turn, may help explain the ecological distribution of this species in Mediterranean-type ecosystems. The experimental results obtained clearly show that the toxic effects of NaCl were much alleviated by the higher light intensity (Table 1). The growth parameters presented by the total dry weight and the leaf number were variant significantly to solar radiation and was affected more in the NaCl-treatment. These results imply that the critical salt-sensitive biochemicals mechanisms affecting plant salt tolerance is affected by solar radiation and those variations can be interpreted as changes in plant salt tolerance. It seemed that the almond tree was more sensitive to salinity than solar irradiation, and this sensitivity was increase by high solar irradiation.
The relation between root and shoot growth emphasizes the dynamic rather than static interdependence of their functional relationships [25,26]. Root-shoot ratios were dependent of rootzone salinity for the both levels of solar radiation investigated. It is a reasonable assumption that plant response in saline environments is a form of optimum autoregulation and that the root/shoot ratio is a state of homeostasis [27], which is independent of root-zone salinity more than solar irradiation. Because the total dry weight is dependent of salinity, it follows from the definition of that the plant can not tolerate higher root-zone salinity mainly at HL by accumulating the rate of salt accumulation relative to its growth rate. Thus, manipulating root/shoot ratios through natural selection or bioengineering might represent an important new strategy to improve plant response in saline and high solar irradiation environments. The observed reduced growth under salt and HL conditions was probably caused by the perturbation of various physiological and biochemical processes at the cellular, tissue or whole-plant level, such as photosynthesis, nutrient uptake and enzymatic activities. In a present work, the reduction in dry weight in plants which experienced was proportional to the reduction in photosynthetic rate, in leaf conductance and transpiration. The results determined that in almond tree signals of chronic photoinhibition were associated with low photosynthetic capacity when the specie was grown under salinity conditions. Nevertheless, it appeared that the A, Gs and E were affected by salinity more then HL treatment and they strongly support the idea that the Mazzetto cultivar examined here actually suffered from salt stress. Our data also provide evidence for the significant interactive effects of sunlight irradiance and salt stress on gaz exchange in almond. We suggest the strong correlation between A and Gs implied that Gs was the dominant limitation to A for all treated plants (Table 2). Stomata closure in response salinity stress generally occurs due to decreased leaf turgor and atmospheric vapor pressure along with root-generated chemical signals [28]. Thus, the decrease in photosynthetic rate under salt-stress conditions is normally attributed to suppression in the mesophyll conductance and the stomata closure at moderate and severe stress [28]. The effects of salinity on photosynthesis are attributed directly to the stomatal limitations for diffusion of gases, which ultimately alters photosynthesis and the mesophyll metabolism [29,28]. The finding of the low effect of sunlight irradiance on photosynthesis rate was actually surprising and contrasts with NaCl effect. It appeared that the reduction of A was correlated negatively to Na+. It seemed that the accumulation of Na+ in leaves depressed the A more than the high light intensity. Thus, Plants grown in the LL intensity treatment were better able to adjust to saline conditions than the plants of the HL treatment. Apparently, the lower light application provided more energy for the regulation of the internal ionic conditions, i.e. for a higher uptake of Ca2+ and Mg2+. This internal ionic regulation probably provided improved conditions for CO2 assimilation and protein synthesis. According to Flowers et al. (1977) [30] internal ionic regulation is a typical response of glycophytes to saline conditions, which obviously depends on light intensity in this present investigation. The effect of light on the Ca2+ content is particularly marked. Several authors have found That Ca2+ is required for maintaining the integrity of plant cell membranes [31]. It thus seems feasible that the depression of lipid formation by NaCl salinity resulted from a lack of Ca2+ under more favourable light conditions it may be argued that more Ca2+ was absorbed by the plants.
Down-regulation of photosynthetic rates is an effective control mechanism for protecting the photosynthetic apparatus from photodamage at low CO2 availability caused by high light irradiation or high salinity [32].
We also observed that, during the treatment period, salt stress acted together with high light irradiance in reducing total chlorophyll and carotenoids content, likely because of the fast imposition of the salt-stress treatment in our experiment, since the same reduction was observed in salt-stressed plants as comparer to unstressed plant in both light levels. However, the decline in both leaf pigments was not the primary cause of the salt-stress-induced reductions in A in high light plants [33]. Leaf Chl content is well established as a common reference system when physiological reactions are quantified [34] and one of the most important factors in determining the photosynthetic rates and dry matter production [35]. The synthesis and the degradation of photosynthetic pigments are normally associated with the photosynthetic efficiency of the plants as well as their growth and adaptability to different environments. The chlorophyll is usually synthesized and photo-oxidised in the presence of light. However, in the current investigation, under conditions of high light intensity, the degradation of chlorophyll molecules might be more intense and equilibrium occurs at low concentration mainly with high level of salinity. It was observed that the chlorophyll concentration in low light leaves treated with NaCl tend to be higher when compared to with NaCl- treated leaves exposed to direct sunlight.
The enhanced chlorophyll content in the low and high light plants under 100 mM NaCl treatment clearly shows that almond could adapt to the level of irradiance by changing chlorophyll contents [36]. Besides the changes in chlorophyll content, change of chloroplast structure also modifies the adaptation of plants under different light conditions interacted with salinity. A much greater quantity of lightharvesting Chl a/b proteins (LHC II) and a fewer number of reaction centres on a total Chl basis affect the high irradiance adaptation response of the photosynthetic pigment apparatus in salt-stressed leaves [37]. Meanwhile, the loss of chlorophyll is often considered as a marker of cellular component of salt stress [38]. Our results clearly show chlorophyll content was reduced by salinity treatments (Table 1).In this study, we suggested that NaCl treatments and the interactions with both level of light had significant effects on the levels of Chl a, Chl b, Chl a+b and Chla/Chlb ratio, but there were not a significant main effects of light. The increase in Chla/Chlb ratio may be due to a conversion of Chlb to Chla. The inter-conversion of Chla and Chlb plays a significant role in the establishment of required Chla/Chlb ratio during the adaptation of stressful conditions [39]. The higher Chla/Chlb ratio indicates also less emphasis on light harvesting in relation to the rates of PSII photochemistry under stress. Increases in Chla/Chlb imply increases in the abundance of entire photosystems and in the size of LHCII, respectively, as virtually all Chla is organized in pigment-protein complexes and LHCII binds most of Chlb [40].
However, it appeared that under high irradiance with high salinity, plants are predisposed to suffer photoinhibition, which is defined as the slow, reversible decline in photosynthetic efficiency that occurs when the absorbed light is in excess of that required for carbon assimilation [41]. This phenomenon is very frequent in plants of the tropics [1], where the light intensity can reach levels over 1800 mol m-2s-1of photosynthetic photon flux density (PPFD) [42].
Because the photosynthetic energy utilization in NaCl treated plants is lower than in unstressed plants, it was assumed that higher antioxidant protection would be necessary to compensate for higher salinity and light-mediated oxidative stress [43]. When plants are exposed to high irradiation associated with a high vapor pressure deficit, the stomata normally close to prevent water loss, which results in a decrease in the intercellular CO concentration and depression of photosynthesis. In this condition, plants become more predisposed to suffer photoinhibition because of the absorption of an excess of photons over what is used in photosynthesis. However, when plants are subjected to such conditions, ROS increases, often resulting in photooxidation [8]. Nevertheless, in such conditions, plants were able to decrease the extent of MDA accumulation in leaves. MDA, a product of lipid peroxidation in plants exposed to adverse environmental conditions, is a reliable indicator of free radical formation and peroxidative damage to cell membranes [8]. Furthermore, MDA has been found to be significantly associated to drought-related oxidative stress in woody plants adapted to arid conditions [44]. The amount of MDA in the leaves was strongly variable in the different treatment studied here. It is worth noting that chlorophyll loss is an efficient photoprotection mechanism that finely modulates the amount of light intercepted by leaves [45]. Therefore, it is not surprising that the highest MDA accumulation observed in 100 mM NaCl-treated plants occurred with lack of chlorophyll loss in LL treatment, and that later on chlorophyll loss in HL of treatment coincided with lower MDA accumulation. In any case, it becomes clear that even when MDA accumulation increases almond plants can withstand this salt-induced oxidative stress, by activating the antioxidant-enzymes systems. Our results demonstrate that almond plants in HL were able to decrease the extent of MDA accumulation in leaves and can withstand doses of salt stress without showing any hint of oxidative stress. This decrease of MDA suggested that membrane peroxidation is not in this case the main detrimental effect caused by high light. However, it is interesting to underline that lower MDA levels in leaves under HL were recorded for NaCl-untreated plants compared to almond plants submitted to NaCl treatment in both level of light treatment. This suggested that almond leaves can contrast oxidative damage. The best protection of almond against oxidative damage could be positively related to an enhanced antioxidant system, especially due to antioxidant enzymes and polyphenols.
Many reports describe detailed changes in the activities of antioxidant enzymes that scavenge ROS in response to high light and salt stress [46]. In our study, in almond tree, the antioxidant enzymes system, including APX, CAT, and POD, responds differently under high light and salt conditions. In this experiment, CAT activity was found to decrease under high sunlight treatment, probably because the enzyme CAT is light-sensitive and suffers from photoinactivation with subsequent degradation in intact leaves as well. Photoinactivation of catalase appears to indicate general symptoms of light sensitivity and early photodamage in leaves [47]. It was postulated that inactivation of CAT could also be mediated by photo-oxidative events initiated through light absorption by chlorophyll [47]. Interestingly, the decreases in CAT activity in both level of light, the main H2O2 -scavenging enzyme, at 100 mM NaCl corresponded to significant decreases in H2O2 levels, so indicating a decrease in H2O2 metabolism at high NaCl levels.
In the antioxidant systems of almond tree, several researchers have reported that higher APX activities protect against environmental stresses such as salt stress [21]. In our study, APX in almond increased by salt stress under LL and by HL, therefore, it remained decrease as response to the interaction effect of salinity and HL. Thus, our results suggest that increased APX activities protect against salt induced oxidative stress when ROS removal is required, such as under LL, but not under HL.
In the present study, the activities of POD rose in all treatment, and this increase was generally higher under salt stress in LL. It appeared that the changes in foliar antioxidant systems correlate more with salinity than light (Table 2), indicating that salinity is an important factor modulating antioxidants [21].
Finally, high sunlight and salinity stress enhance the biosynthesis of polyphenols, particularly flavonoids, likely using ROS as signalling molecules [11]. Phenolics are the main class of secondary metabolites with important roles in abiotic stress tolerance due to their antioxidant properties [48]. In this present investigation, it appeared that the salt and light induced polyphenols content in leaves of Mazzetto, but this increase was higher under the combined effect of salt and HL. We suggested that the interaction of salt stress and the HL increase the photoproctection demand of almond tree by inducing the polyphenols pathways quite the reverse effect of antioxidant enzymes.
References
- Krause GH, Koroleva OY, Dalling JW, Winter K (2001) Acclimation of tropical tree seedlings to excessive light in simulated tree-fall gaps. Plant Cell and Environment 24: 1345-1352.
- Demmig-Adams B, William W A (1993) The Xanthophyll Cycle, Protein Turnover, and the Tolerance of Sun-Acclimated Leaves. Plant Physiol 103: 1413-1420.
- Mittler R, Vandarauwera S, Gollery M, Van Breusegem F (2004) Reactive oxygen network of plants. Trends Plant Sci 9: 490-498.
- 4. Ort RD, Baker NR (2002) A photoprotective role for O2 as an alternative electronic sink in photosynthesis? Curr Opin Plant Biol. 5: 193-198.
- Halliwell B (2006) Reactive Species, Antioxidants, Redox biology is a fundamental theme of aerobic life. Plant Physiol 141: 312-322.
- Mittler R (2006) Abiotic stress, the field environment and stress combination. Trends Plant Sci 11: 15-19.
- Lu C, Jiang G, Wang B, Zhang J (2003) Salinity treatment shows no effects on photosystem II photochemistry but increases the resistance of photosystem II to heat stress in the halophyte Suaeda salsa. J Exp Bot 59: 851-860.
- Asada K (2006) Production and scavenging of reactive oxygen Species in chloroplasts and their functions. Plant Physiol 141: 391-396.
- Muller P, Xiao-Ping L, Niyogi K K (2001) Non-photochemical quenching. A response to excess light energy. Plant Physiol 125: 1558-1566.
- Peterman EJG, Dukker FM, Van Grondelle R, Van Amerongen H (1995) Chlorophyll a and carotenoid triplet states in light-harvesting complex II of higher plants. Biophys J 69: 2670-2678.
- Tattini M, Remorini D, Pinelli P, Agati G, Saracini E, et al. (2006) Morpho-anatomical, physiological and biochemical adjustments in response to root zone salinity stress and high solar radiation in two Mediterranean evergreen shrubs, Myrtus communis and Pistacia lentiscus. New Phytol 170: 779-794.
- Tattini M Traversi M L (2008) Responses to changes in Ca2+supply in two Mediterranean evergreens, Phillyrea latifolia and Pistacia lentiscus, during salinity stress and subsequent relief. Ann Bot 102: 609-622.
- Caldwell MM, Bornman JF, Ballare CL, Flint SD, Kulandaivelu G (2007) Terrestrial ecosystems, increased solar ultraviolet radiation, and interactions with other climate change factors. Photochem Photobiol Sci 6: 252-266.
- Agati G, Matteini P, Goti A, Tattini M (2007) Chloroplast-located flavonoids can scavenge singlet oxygen. New Phytol 174: 77-89.
- Foyer CH, Lelandais M, Kunert KJ (1994) Photooxidative stress in plants. Plant Physiol 92: 696-717.
- 16. Faveretto FV, Martinez CA, Soriani HH, Furriel RPM (2011) Differential responses of antioxidant enzymes in pioneer and late successional tropical tree species grown under sun and shade conditions. Environ Exper Bot 70: 20-28.
- Munns R, James RA (2003) Screening methods for salinity tolerance: a case study with tetraploid wheat. Plant Soil 253: 201-218.
- Arno DI (1949) Copper enzyme in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol 24: 1-15.
- Mınguez-Mosquera MI, Hornero-Mendez D (1993) Separation and quantification of the carotenoid pigments in red peppers (Capsicum annuum L.), paprika, and oleoresin by reversed phase HPLC. J Agric Food Chem 41: 1616-1620.
- Tomás-Barberán FA, Gil MI, Cremin P, Waterhouse AL, Hess-Pierce B, Kader AA (2001) HPLD-DAD-ESIMS analysis of phenolic compound s in nectarines, peaches, and plums. J Agric Food Chem 49: 4748-4760.
- Sorkheh K, Shiran B, Rouhi V, Khodambashi M, Sofo A (2011) Regulation of the ascorbate-glutathione cycle in wild almond during drought stress. Russ J Plant Physiol 58: 76-84.
- Hodges DM, DeLong JM, Forney CF, Prange RK (1999) Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta. 207: 604-611.
- Nakano Y, Asada K (1981) Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol 22: 867-880.
- Osswald WF, Kraus R, Hippeli S, Benz B, Volpert R, et al. (1992) Comparison of the enzymatic activities of dehydroascorbic acid reductase, glutathione reductase, catalase peroxidase and superoxide dismutase of healthy and damaged spruce needles (Picea abies L.). J Plant Physiol 139: 742-748.
- Baker JM, Wraith JM, Dalton FN (1991) Root function in water transport. Adv Soil Sci 19: 54-67.
- Dalton FN (1996) In-situ extent measurements by electrical capacitance methods. Plant Soil 173: 157-165.
- Klepper B (1991) Root Shoot Relationships. In Plant Roots, The Hidden Half. Eds. Y. Waisel, A. Eshel and U. Kafkafi. Marcel Dekker, Inc. New York. 948 p.
- Chaves MM, Zarrouk O, Francisco R, Costa JM, Santos T, et al. (2009) Grapevine under deficit irrigation: hints from physiological and molecular data. Ann Botany 105: 661-676.
- Parida A, Chattopadhyay G, Kumar U (2005) Multi criteria maintenance performance measurement: a conceptual model. Proceedings of the 18th International Congress COMADEM, 31st Aug-2nd Sep 2005, Cranfield, UK, pp 349-356.
- Flowers TJ, Troke PF, Yeo AR (1977) The mechanism of salt tolerance in halophytes. Annu Rev of Plant Physiol l28: 89-121.
- Van Steveninck RKM (1965) The significance of calcium on the apparent permeability of cell membranes and the effect of substitution with other divalentions. Physiol Plant 18: 54-69.
- Chaves MM, Pereira JS, Maroco J, Rodrigues ML, Ri cardo CPP, et al. (2002) How plants cope with water stress in the field. Photosynthesis and growth. Ann Botany 89: 907-916.
- Flexas J, Bota J, Galmos J, Medrano H, Ribas-Carb M (2006) Keeping a positive carbon balance under adverse conditions: responses of photosynthesis and respiration to water stress. Plant Physiol 127: 343-352.
- Wittmann C, Aschan G, Pfanz H (2001) Leaf and twig photosynthesis of young been (Fagus sylvstica) and aspen (Populus tremula) trees grown under different light regime. Basic Appm Ecol 2: 145-154.
- Mao LZ, Lu HF, Wang Q, Cai MM (2007) Comparative photosynthesis characteristics of Calycanthus chinensis and Chimonanthus praecox. Photosynthetica 45: 601-605.
- Dai YJ, Shen ZG, Liu Y, Wang LL, Hannawayc D, Lu HF (2009) Effects of shade treatments on the photosynthetic capacity, chlorophyll fluorescence, and chlorophyll content of Tetrastigma hemsleyanum Diels et Gilg. Environ Exper Bot 65: 177-182.
- Laisk A, Eichelmann H, Oja V, Rasulov B, Padu E, et al. (2005) Adjustment of leaf photosynthesis to shade in a natural canopy: rate parameters. Plant Cell and Environment 28: 375-388.
- Singh AK, Dubey R S (1995) Changes in chlorophyll a and b contents and activities of photosystems I and II in rice seedlings inducedby NaCl. Photosynthetica 31: 489-499.
- Mane AV, Karadge BA Samant JS (2010) Salinity induced changes in photosynthetic pigments and polyphenols of Cymbopogon nardus (L.) Rendle. J Chem Phys Rese 2: 38-347.
- Anderson JM, Chow WS, Park YI (1995) The grand design of photosynthesis: acclimation of the photosynthetic apparatus to environmental cues. Photosynth Res 46: 129-139.
- Long SP, Humphries S, Falkowski PG (1994) Photoinhibition of photosynthesis in Nature. Annu Rev Plant Physiol Plant Mol Biol 45: 633-662.
- Larcher W (1995) Physiological Plant Ecology. Springer, Berlin, 506 pp.
- Hansen LD, Churchb JN, Mathesona S, McCarlie VW, Thygersonc T, et al. (2002) Kinetics of plant growth and metabolism. Thermochim Acta 388: 415-425.
- Bacelar EA, Santos DL, Moutinho-Pereira JM, Lopes JI, Goncalves BC, et al. (2007) Physiological behaviour, oxidative damage and antiox-idative protection of olive trees grown under different irrigation regimes. Plant Soil 292: 1-12.
- Munne-Bosch S, Alegre L (2002) Plant aging increases oxidative stress in chloroplasts. Planta 214: 605-615.
- Ma F, Cheng L (2003) The sun-exposed peel of apple fruit has higher xanthophyll cycle- dependent thermal dissipation and antioxidants of the ascorbate glutathione pathway than the shaded peel. Plant Sci. 165: 819-827.
- Feierabend J, Schaan C, Hertwig B (1992) Photoinactivation of catalase occurs under both high- and low-temperature stress conditions and accompanies photoinhibition of photosystem II. Plant Physiol 100: 1554-1561.
- Ruiz J M, Romero L (2001) Bioactivity of the phenolic compounds in higher plants. In: Atta-ur-Rahman (ed) Studies in natural products chemistry. Bioactive natural products, 25. part F. Elsevier, Oxford, pp 651-683.